При встрече фага и чувствительных к нему бактерий наблюдается несколько последовательных стадий, в результате которых происходит разрушение бактерий или их лизис.
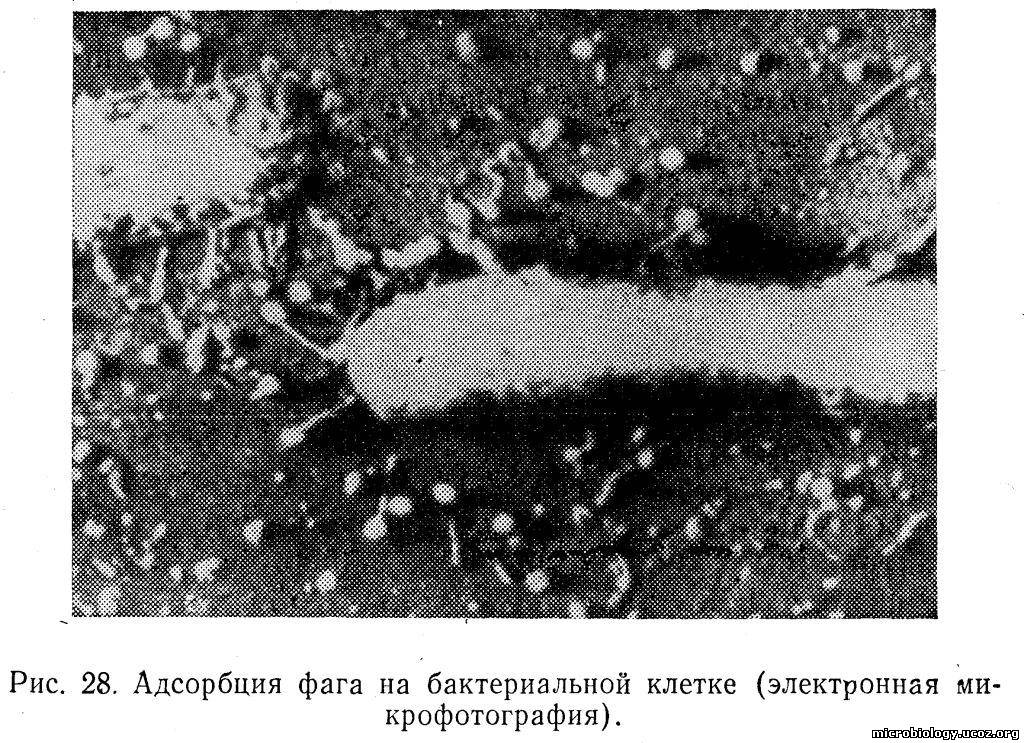
Адсорбция фага заключается в фиксации корпускул фага на специальных фаговых рецепторах, расположенных на клеточной стенке бактерий (рис. 28).
После того как отросток фага прикрепился к специфическому рецептору клетки своими нитями, жесткая трубка цилиндра прокалывает клеточную стенку бактерии. В цитоплазматической мембране клетки образуется отверстие под действием фермента отростка, сходного с лизоцимом. Через это отверстие ДНК, содержащаяся в головке фага, впрыскивается через канал отростка в цитоплазму бактерии. Пустая оболочка фага в виде «тени» остается снаружи клетки. Количество фаговых частиц, которые могут быть адсорбированы бактериальной клеткой обусловлено площадью ее адсорбционной поверхности и величиной фага. Например, 1/3 поверхности бактерии размером 1*3 мкм может адсорбировать 300 частиц фага величиной около 100 нм.
Латентный период наступает после проникновения ДНК фага в цитоплазму бактерии. Длительность этого периода зависит от системы фаг — хозяин. Так, для кишечной палочки и ее бактериофага Т2 он составляет 12 мин. В этот период фаг находится в скрытом, латентном состоянии, и, если даже разрушить клетку, то ДНК фага обнаружить невозможно.
Период образования нового фага представляет собой очень сложный процесс, во время которого фаговая ДНК с невероятной быстротой «захватывает власть» в бактериальной клетке, подавляя ее синтезирующие механизмы. ДНК фага принимает на себя генетическое управление жизнедеятельностью бактерий, которые в своих рибосомах начинают синтезировать белки фага и фаговые оболочки. По матрице ДНК фага реплицируются, создаются ее молекулы. В конце латентного периода фаговые ДНК и белковые оболочки соединяются в цитоплазме бактерии и образуют зрелые частицы фага.
Лизис клетки наступает приблизительно через 30 мин после прикрепления фага к поверхности бактерии. Полагают, что лизис вызывается или зависит от образования специального литического фермента, сходного с лизоцимом. Образовавшиеся корпускулы фага освобождаются и начинают новый цикл развития в других бактериальных клетках. Число вирусных частиц, образующихся в каждой инфицированной бактериальной клетке, примерно постоянно для определенной системы фаг — бактерия: от 20 до 200 и более. Это число называют выходом фага. Фаги, вызывающие лизис бактериальных клеток, называют вирулентными. В отличие от них существуют фаги, которые, проникая в бактерии, не разрушают их, а длительно в латентном состоянии присутствуют в них, размножаясь вместе с бактериальной клеткой. Такие фаги называют умеренными.