Покрытосеменные, наиболее молодая группа растений, быстро распространились, начиная с середины мезозойской эры, и занимают в настоящее время господствующее положение среди растений земного шара.
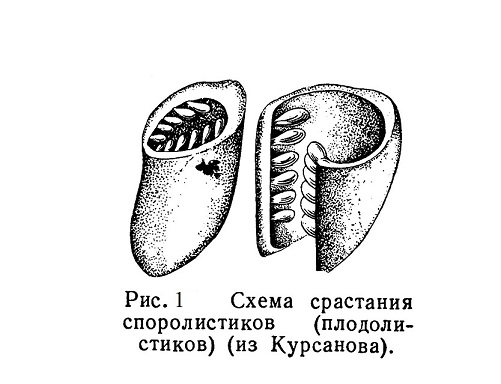
По сравнению с голосеменными главным новшеством у покрытосеменных является возникновение пестика, образовавшегося из свернувшихся и сросшихся краями плодолистиков (рис. 1). Расположенные на споролистиках макроспорангии с покровами (семяпочки) заключены в образованную таким путем полость.
Семя развивается под «прикрытием» сросшихся споролистиков (отсюда название подтипа — покрытосеменные) (рис. 1).
В пестике различают:
1) нижнюю расширенную часть — завязь, в которой после оплодотворения развивается — «завязывается» — семя. Семя возникает из семяпочки (макроспорангия с покровом), как мы это видели и у голосеменных; плод, заключающий в себе семена, является новообразованием и развивается из завязи — нижней части сросшихся споролистиков, которые поэтому здесь называются плодолистиками;
2) над завязью находится суженная часть пестика — столбик, несущий на своей верхушке
3) рыльце (рис. 2).
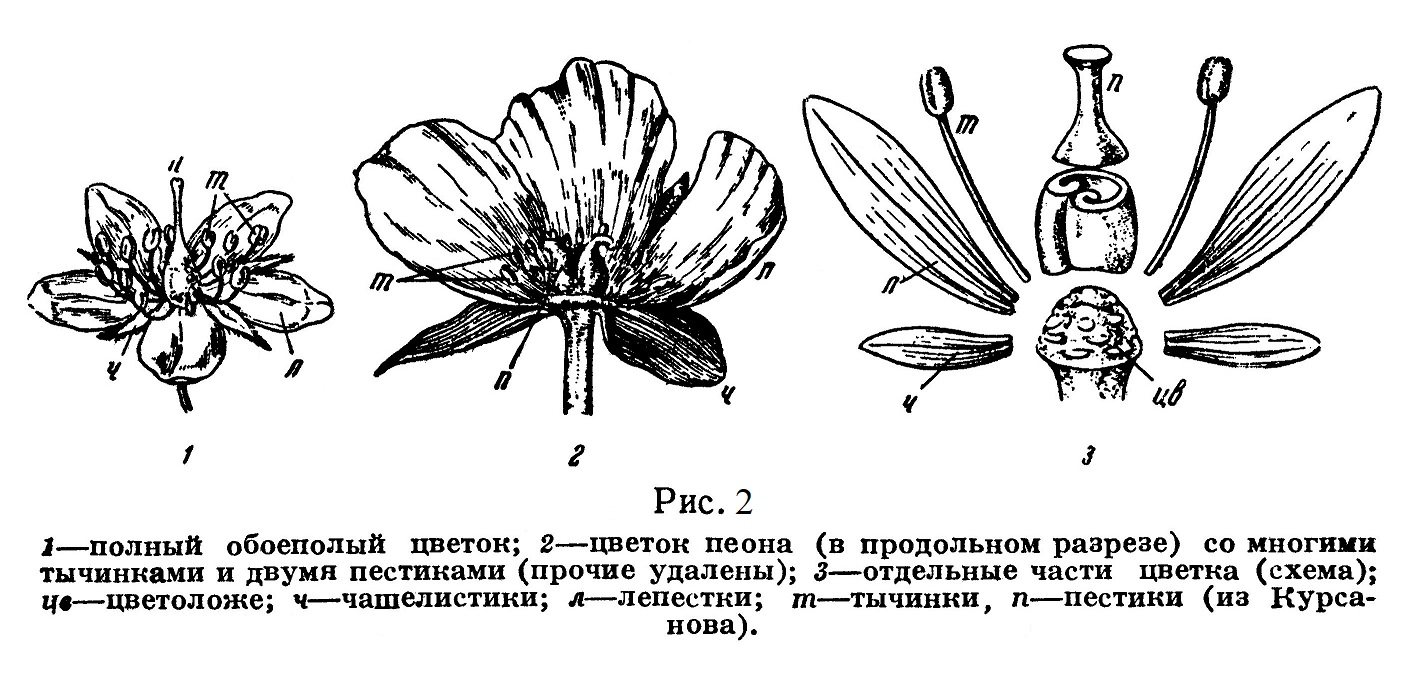
Таким образом, развивающиеся из семяпочек семена покрытосеменных защищаются не только покровами, впервые появившимися еще у голосеменных, но и новым образованием — пестиком, завязь которого после оплодотворения развивается в плод.
Микроспоролистики покрытосеменных преобразовываются в тычинки (рис. 2), обычно состоящие из двух частей — из тычиночной нити и пыльника, большей частью четырехгнездного, представляющего собой сравнительно мало измененные парные микроспорангии; в них развиваются микроспоры — пылинки или пыльцевые зерна.
Пестики тычинки, обычно называемые половыми органами, являются, как видно из истории их происхождения, сильно измененными листьями спорофита — макро- и микроспоролистиками, органами бесполого размножения.
Однако в развивающихся на них макро- и микроспорах уже заложены биологические особенности будущего полового поколения.
Одним из морфологических проявлений этих особенностей спор и развивающегося из них гаметофита является гаплоидный набор хромосом, как и у всех описанных листостебельных растений.
Кроме возникновения пестика и развивающегося из него плода, покрытосеменные характеризуются появлением цветка, откуда второе название этого подтипа — цветковые растения.
В отличие от шишки голосеменных цветок представляет собой, кроме скученных споролистиков, также присоединенные к ним группы других видоизмененных листьев, играющих в процессе эволюции разнообразную роль и составляющих так называемый околоцветник. Последний обычно состоит из наружной части — чашечки и внутренней — венчика, образованного часто разнообразно окрашенными лепестками (рис. 2).
Как ни видоизменены части цветка, мы и у современных растений видим доказательства их листового происхождения.
Наименее изменены чашелистики, составляющие самую наружную часть околоцветника — чашечку. Отличаясь от вегетативных листьев формой и размерами, чашелистики сохраняют основную особенность листа — зеленую окраску. На некоторых цветках можно легко проследить ряд переходов от чашелистиков к лепесткам, еще более измененным листьям, и затем далее — от лепестков к тычинкам.
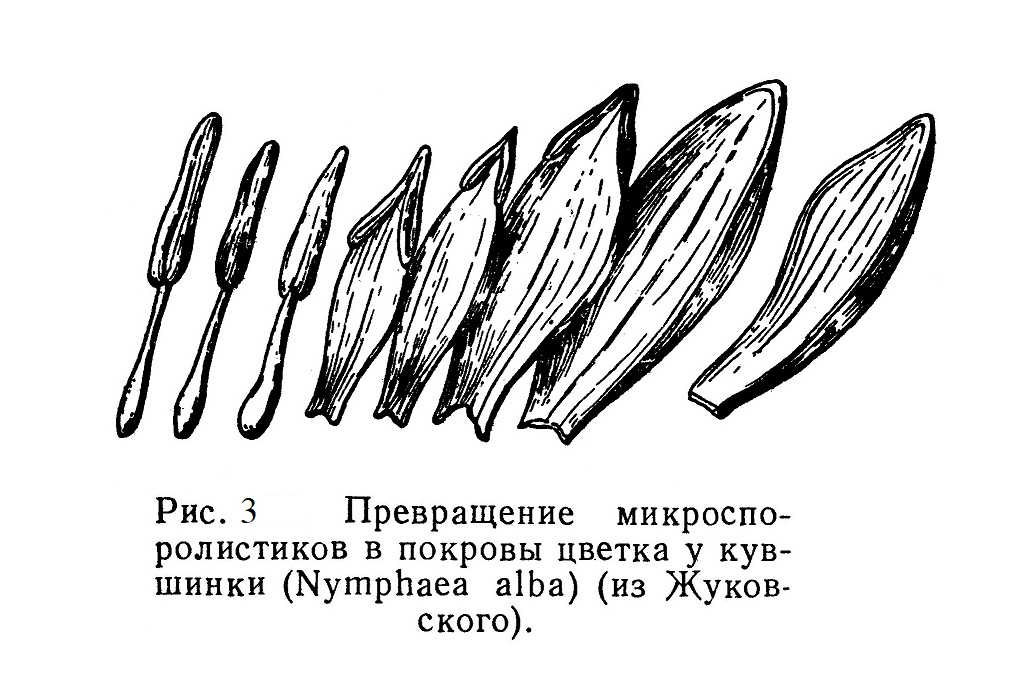
В цветах белой кувшинки мы видим на чашелистиках — зеленых снаружи и белых изнутри — переход к белым лепесткам. Еще дальше внутрь видно появление пыльников на белых лепестках; последние по направлению к центру цветка постепенно суживаются и превращаются в белые тычиночные нити (рис. 3).
Махровые цветы (розы, тюльпаны, гвоздики и др.) — пример обратного перехода тычинок и пестиков в лепестки (рис. 4).

Итак, цветок представляет собой видоизмененный стебель с видоизмененными листьями (видоизмененный побег). Укороченная и расширенная верхушка стебля утратила способность к неограниченному росту и превратилась в расширенную часть, несущую все органы цветка — цветоложе.
Некоторый свет на происхождение покрытосеменных проливают скудные палеонтологические остатки бенеттитов.
Напоминая внешне древовидные папоротники и саговники (рис. 5), бенеттиты характеризуются наличием органов размножения, близко стоящих к современным цветкам (рис. 6). Мы встречаем у них впервые собранные вместе группы макроспорофиллов и микроспорофиллов, причем последние окружают первые.

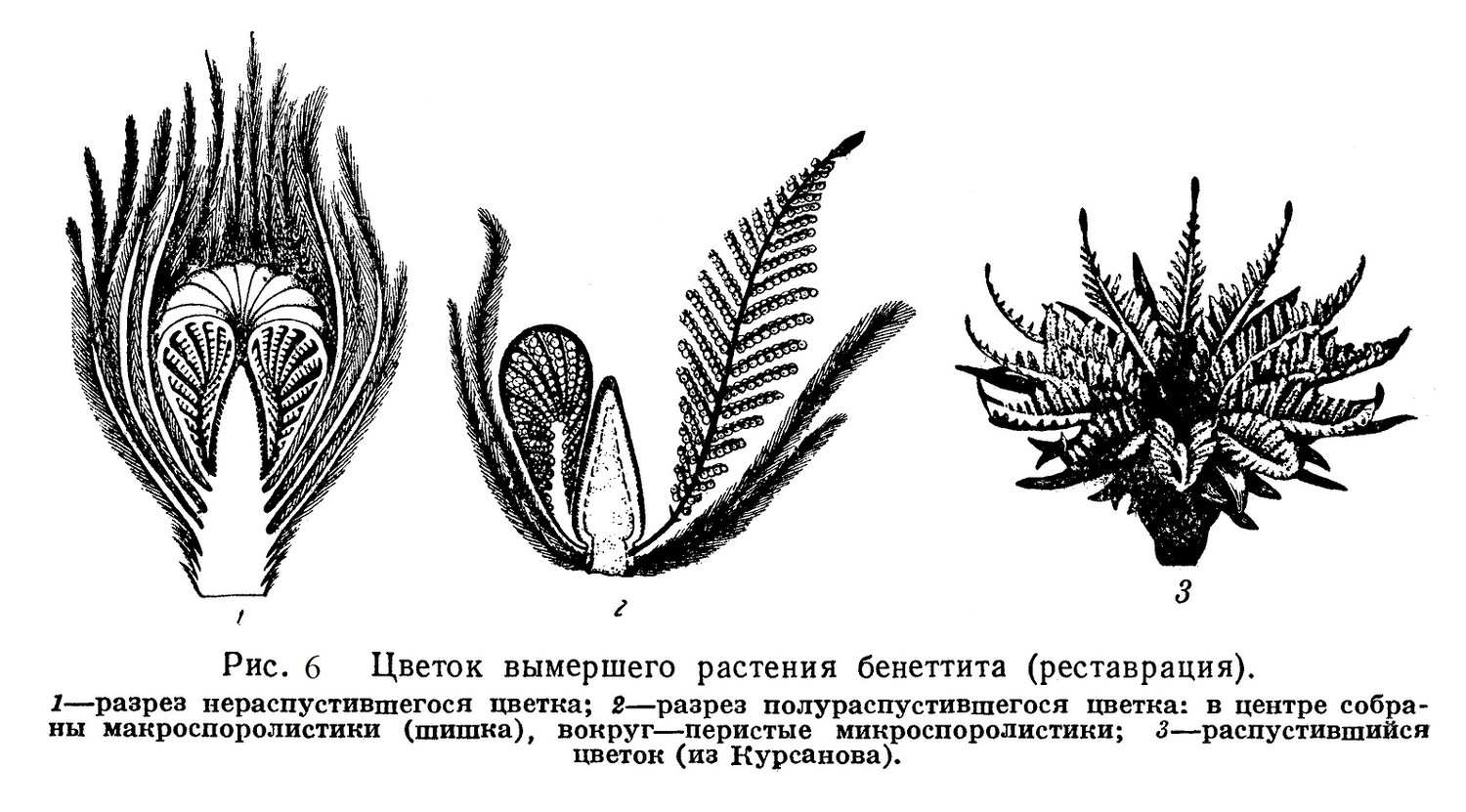
Тесное сближение и взаимное расположение тех и других споролистиков напоминают расположение соответствующих органов в современных «обоеполых» цветках. Кроме того, в цветках бенеттитов, наряду с группами споролистиков, имелись также и бесплодные листки, игравшие, возможно, защитную роль и являвшиеся, по-видимому, предшественниками околоцветника.
Наряду с этими прогрессивными чертами, некоторые бенеттиты несут и примитивные особенности — их микроспоролистики еще очень напоминают своей перисторассеченной формой вегетативные листья папоротников и др.
Бенеттиты появились в начале средней (мезозойской) эры и просуществовали до конца ее, наряду с возникшими в средине мезозойской эры настоящими покрытосеменными. Многие систематики причисляют их к голосеменным на основании описанных выше черт, но считают их в то же время организмами, переходными к покрытосеменным.
Среди современных цветов чрезвычайно распространено явление «гермафродитизма» — наличие в одном цветке и макро- и микроспоролистиков.
Согласно изложенному взгляду на «цветок» бенеттитов, такие «обоеполые» цветы являются более древними образованиями. Раздельнополость цветов — явление более позднее.
В настоящее время однополые цветы менее распространены в природе, чем двуполые, и возникли, может быть, в процессе эволюции как одно из приспособлений к перекрестному опылению.
Это соображение относится в особенности к так называемым двудомным растениям, у которых однополые цветы разных полов размещены на разных экземплярах, например, у ив, тополя, конопли и др. Однодомными называются растения, несущие также однополые цветы, однако расположенные на одном и том же экземпляре растения, например, у кукурузы, березы, орешника, дуба, тыквы, огурцов.
Как известно, близкородственное скрещивание ведет к обеднению наследственной основы организма (Лысенко), к снижению его жизнеспособности. Еще Дарвин посвятил в связи с этим много труда изучению разнообразных, часто удивительно тонких приспособлений к перекрестному опылению, выработавшихся путем естественного отбора в истории цветка.
Перекрестное опыление заключается в перенесении пыльцы (микроспор) на макроспоролистик — на пестик другого цветка, точнее — на его рыльце.
Перенос этот совершается при помощи либо неживой природы (ветер, реже течение воды), либо при помощи животных.
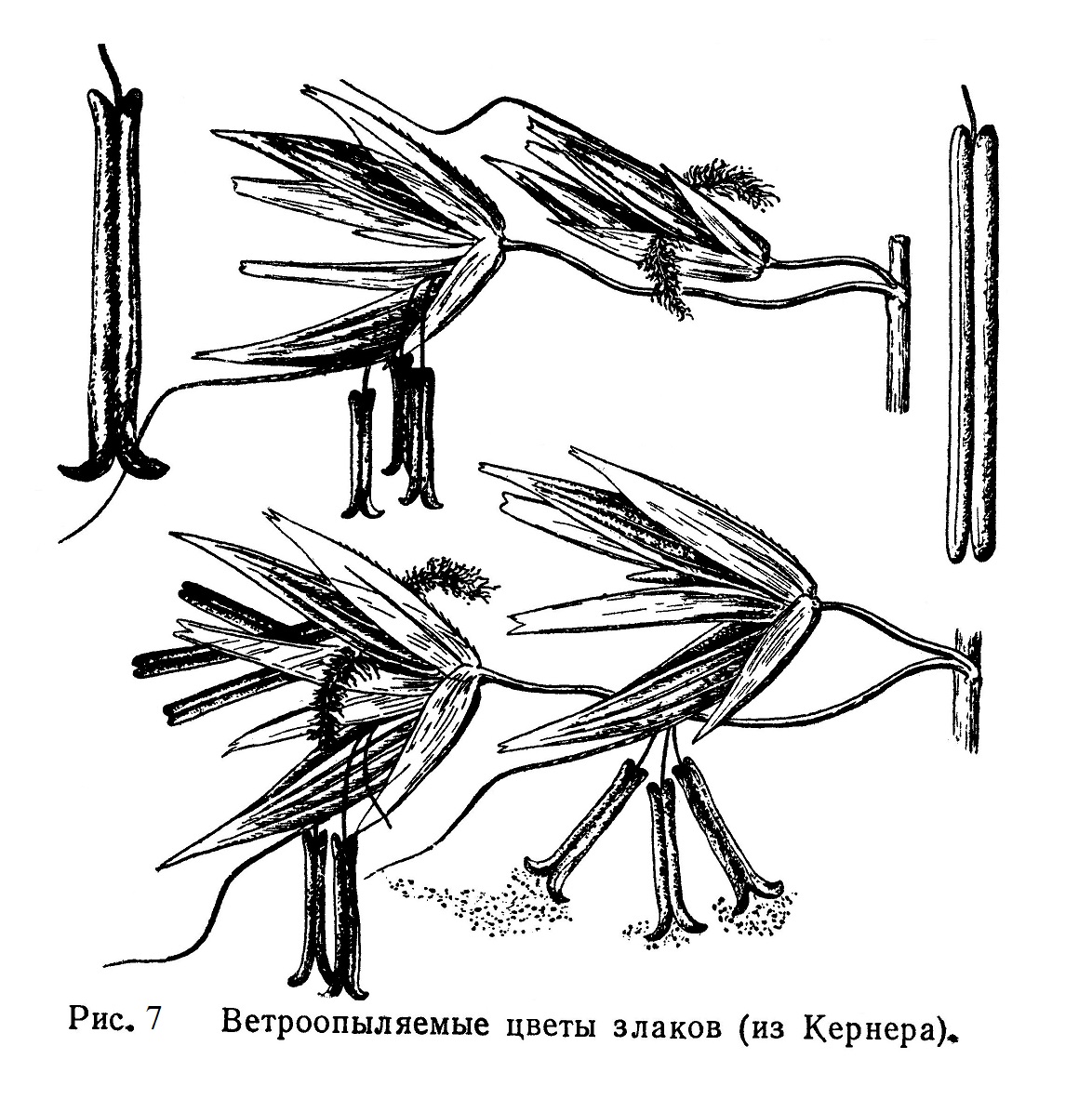
Характерными особенностями ветроопыляемых цветов (анемофильных) являются обильная продукция пыльцы, ее легкость и сухость, крупное, часто торчащее из цветка рыльце пестика, улавливающее летящую пыльцу, и отсутствие крупного околоцветника, который мог бы мешать ветроопылению. Такие цветы невзрачны и не привлекают нашего внимания (например, цветы злаков, рис. 7).
Совсем другие черты мы видим у цветов, опыляемых при помощи животных, главным образом при помощи насекомых.
В процессе длительной эволюции покрытосеменных — цветковых — растений выработался ряд черт, связывающих растения с их опылителями. Околоцветник, игравший первоначально защитную роль, приобретает в течение эволюции разнообразную окраску, резко выделяющуюся на фоне зелени листвы; цветы привлекают к себе насекомых (бабочек, пчел, шмелей, двукрылых и др.), а иногда и мелких птиц (колибри); некоторые из них питаются либо сладким нектаром, либо избытком пыльцы. Железки, вырабатывающие сладкий нектар, — нектарники — являются одной из характерных черт насекомоопыляемых (энтомофильных) цветов (рис. 8).
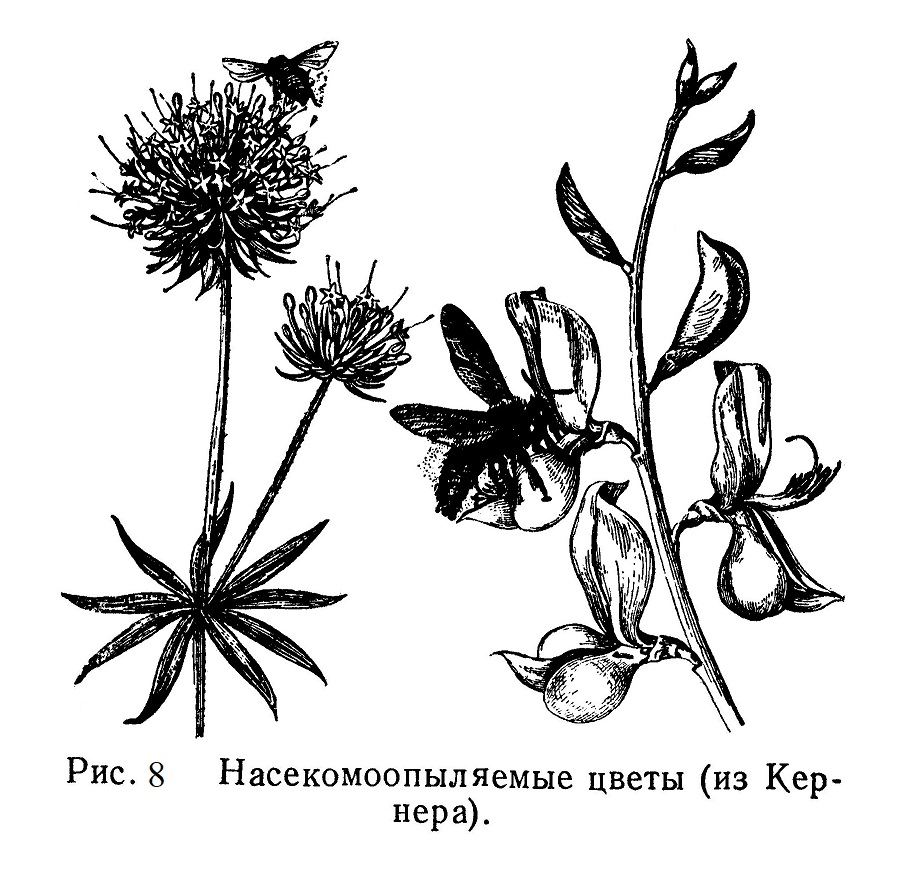
Глаз насекомых привлекает не только окраска, но и размер и подчас причудливая форма цветов. В связи с этим мы видим часто сгруппированные мелкие цветы — так называемые соцветия, в глаза не меньше, чем крупные одиночные цветы (рис. 9).

Одним из весьма распространенных способов привлечения насекомых цветами являются разнообразные запахи — от нежных ароматов душистых цветов, привлекающих пчел, бабочек и других насекомых, до запаха падали и гниения, привлекающего разные виды мух. Путем естественного отбора создались весьма разнообразные приспособления к перекрестному опылению при посредстве насекомых. Особенно тонкие приспособления к перекрестному опылению имеются у обоеполых цветов.
Посещая определенные цветы как источник питания, насекомые в свою очередь часто очень тонко приспособлены к определенным видам цветов (устройство хоботка, лапок и пр.). Пыльца прилипает к разным частям тела насекомого и переносится им с одного цветка на другой тога же вида растений.
В связи с этим пыльца насекомоопыляемых растений обладает иными особенностями, чем пыльца ветроопыляемых: она липкая, часто снабженная шипиками, выростами оболочки и другими особенностями, облегчающими прикрепление их к посещающему цветок насекомому.
Все описанные до сих пор черты покрытосеменных растений касаются спорофита высокоорганизованного листостебельного растения, на котором диференцируются разнообразно видоизмененные листья, связанные с размножением путем продукции макро- и микроспор. Как и у голосеменных, макроспора прорастает внутри макроспорангия, из которого она не выходит, оставаясь вместе с женским гаметофитом на материнском растении.
Как и у голосеменных, микроспора прорастает еще в микроспорангии, в который она затем покидает, переносясь к макроспоролистикам вместе с мужским гаметофитом, не выходящим из микроспоры.
Однако между прорастанием спор у покрытосеменных и у голосеменных имеются и существенные отличия, особенно в отношении женского гаметофита. Последний у покрытосеменных редуцирован значительно больше, чем у голосеменных.
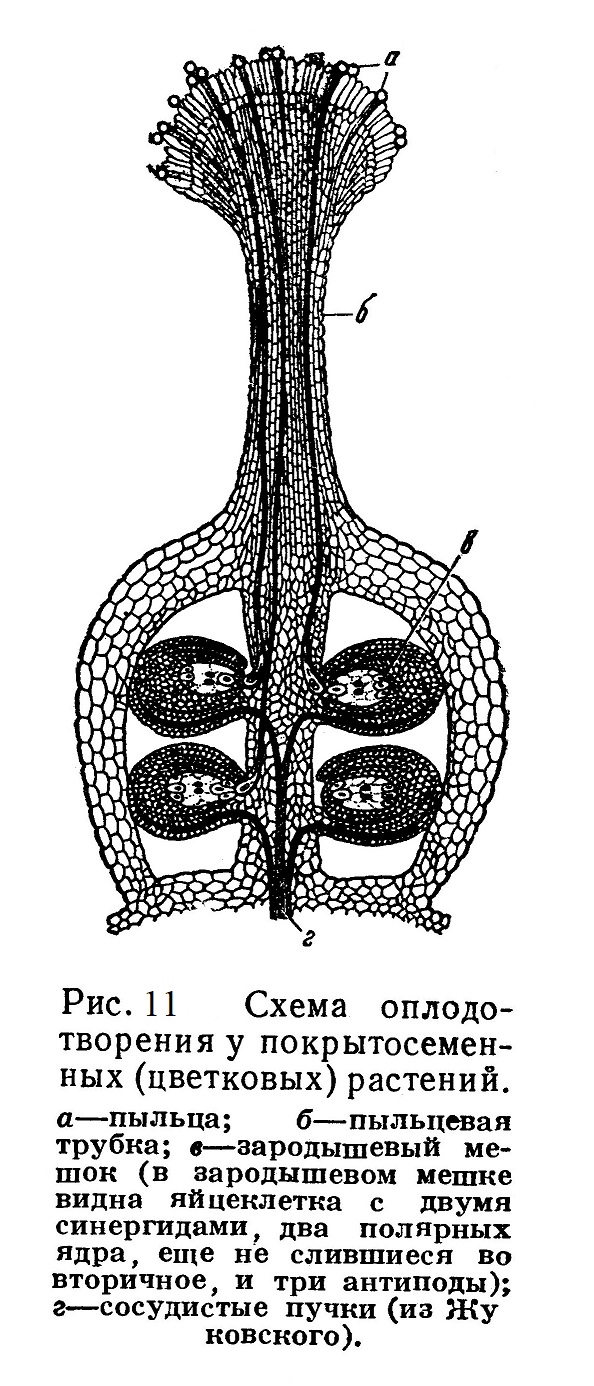
При прорастании макроспоры предварительно трижды происходит деление ее ядра; из образовавшихся 8 ядер три располагаются на одном полюсе макроспоры — зародышевого мешка, три — на другом полюсе; вокруг каждого из них обособляются участки цитоплазмы, и таким образом на каждом полюсе формируется по 3 клетки. Два из 8 ядер сливаются, образуя вторичное ядро крупной центральной клетки зародышевого мешка (рис. 12). Одна из трех клеток, находящаяся ближе к пыльцевходу, представляет собой яйцеклетку; остальные две — синергиды (спутницы); на противоположном полюсе расположены антиподы.
Полученные в итоге 7 клеток, лежащие внутри общей оболочки макроспоры (зародышевого мешка), представляют собой весьма редуцированный женский гаметофит. Архегониев уже нет.
Мужской гаметофит развивается внутри микроспоры (пыльцевого зерна), представляющей собой первоначально, как и макроспора, одну клетку. При развитии мужского гаметофита микроспора делится на две клетки — крупную, вегетативную клетку и лежащую в ней веретеновидную генеративную (рис. 10, А). Последняя также делится, давая два спермия — мужские половые клетки, лишенные, как и у хвойных, органоидов движения (рис. 10, Б, В).
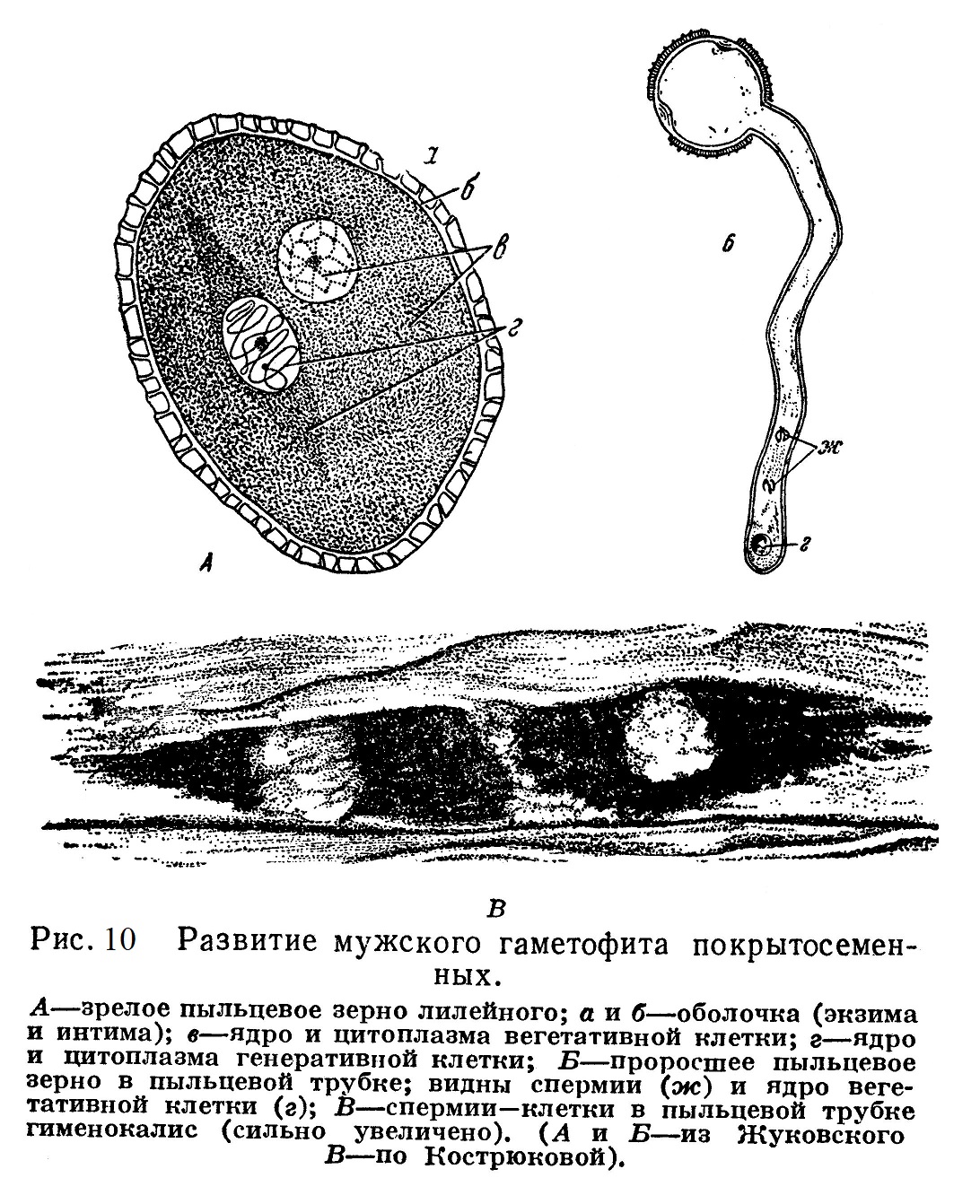
Зрелая пылинка переходит в покоящееся состояние, которое наступает у разных растений на разных этапах развития мужского гаметофита. В связи с этим деление генеративной клетки часто происходит позже.
Попав на рыльце пестика, выделяющее сахаристые вещества, вегетативная клетка пылинки прорастает в длинную пыльцевую трубочку, врастающую в канал столбика и дорастающую до семяпочек (макроспорангиев), лежащих в завязи (рис. 11).
У некоторых растений деление генеративной клетки происходит уже в растущей пыльцевой трубке, по которой они передвигаются в направлении ее роста (рис. 10). После проникновения в зародышевый мешок оболочка пыльцевой трубочки растворяется й спермин попадают в зародышевый мешок.
Оба проникшие в зародышевый мешок спермия участвуют в оплодотворении: один из них сливается с яйцеклеткой, давая начало развивающемуся из зиготы зародышу нового спорофита; другой спермий сливается с центральной клеткой зародышевого мешка, содержащей вторичное ядро {рис. 12). В результате этого второго оплодотворения также происходит деление клеток и развивается многоклеточная ткань — так называемый эндосперм, в которой накопляется запас питательных веществ; за счет этих питательных веществ впоследствии идет рост зародыша на первых порах его самостоятельной жизни.
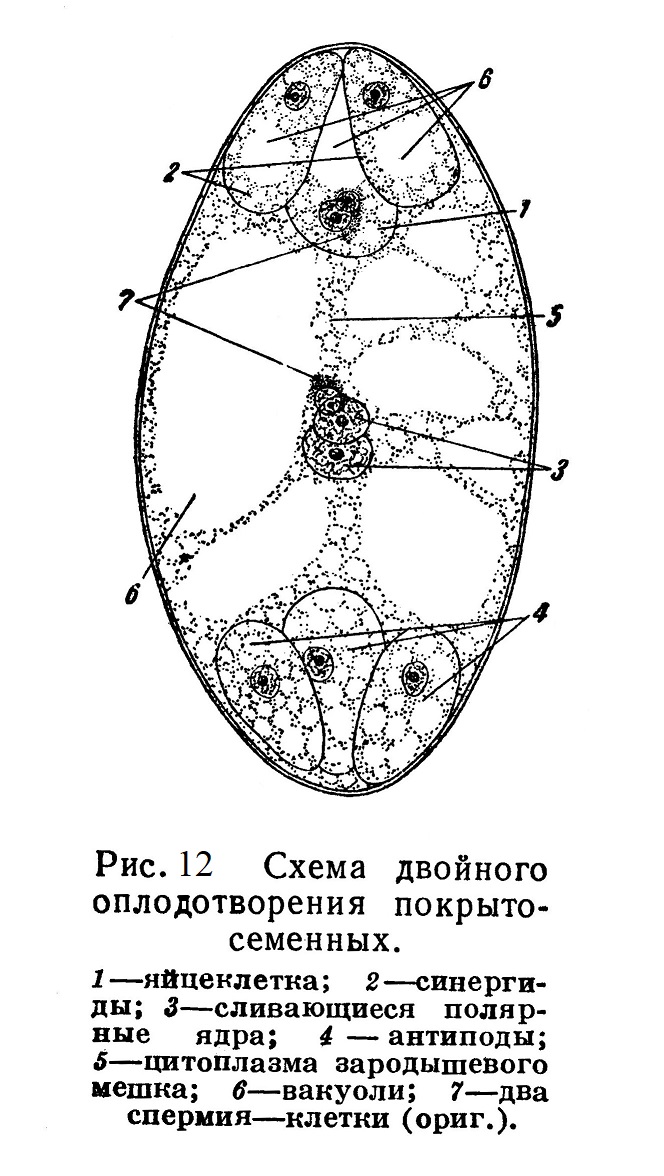
Описанное явление носит название двойного оплодотворения. Оно было открыто профессором Киевского университета С. Г. Навашиным и является существенным новообразованием, присущим только покрыто-семенным.
Если у голосеменных питание зародыша идет вначале за счет пита-тельных веществ самого женского гаметофита, то у цветковых растений питающая ткань, как и сам зародыш, возникает в результате оплодотворения; следовательно, не только зародыш, но и пища его несет в себе двойственную наследственность, что повышает жизненность нового поколения.
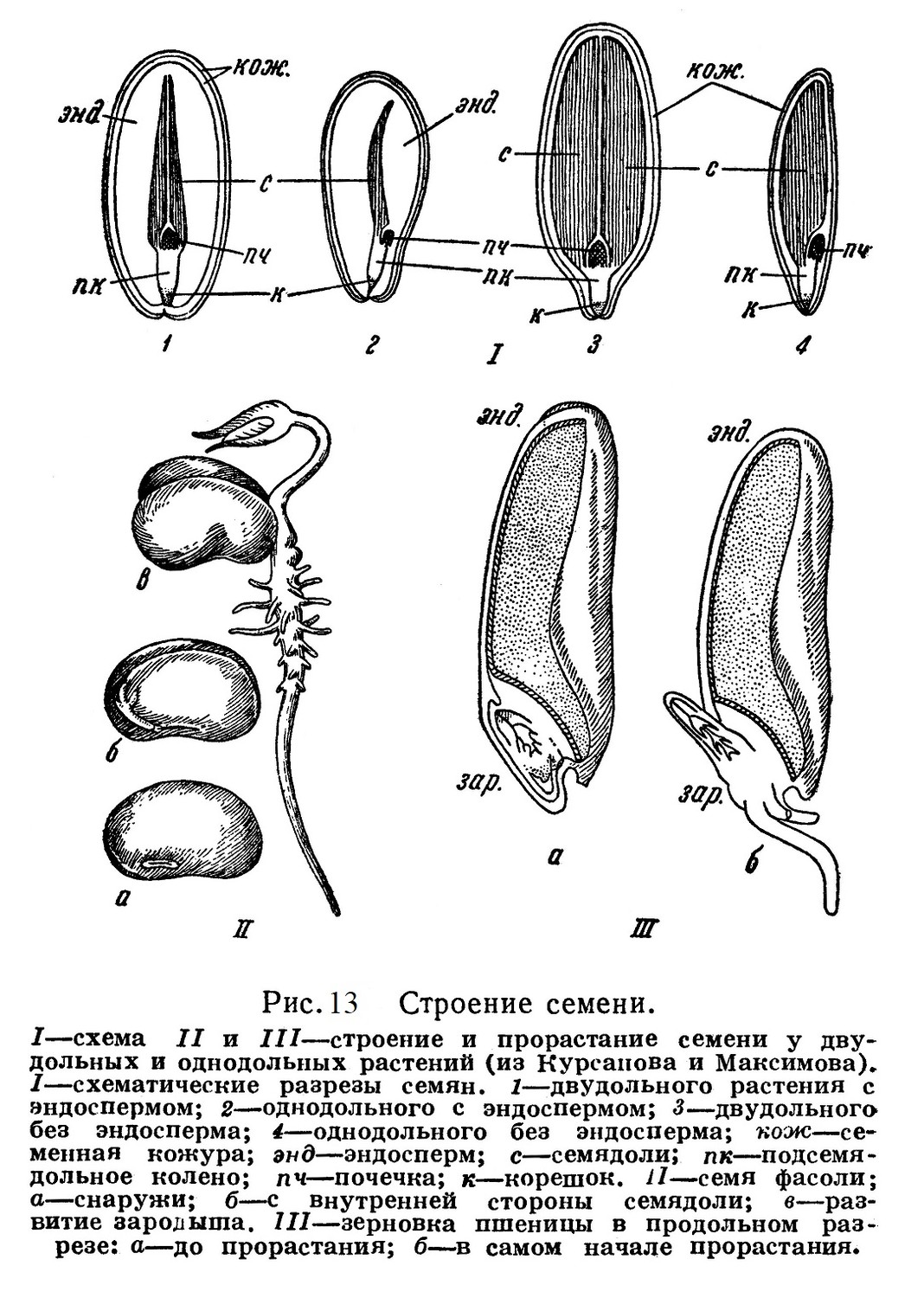
Итак, семя покрытосеменных состоит из зародыша и эндосперма причем в отличие от голосеменных не только зародыш, но и эндосперм возникает в результате оплодотворения. Снаружи семя покрыто плотной защитной кожурой, развивающейся, как и у голосеменных, из покровов семяпочки — макроспорангия.
У некоторых растений зародыш еще во время формирования семени потребляет весь запас питательных веществ эндосперма и откладывает в себе самом новые, им самим переработанные питательные вещества.
Откладываются эти запасы в таких случаях в видоизмененных листочках зародыша — в его семядолях. Такое явление наблюдается в семенах бобовых (фасоль, горох и др.), подсолнечника и др. (рис. 13).
Долгие годы шел спор о том, являются ли спермин покрытосеменных клетками или ядрами. Убеждение в отсутствии цитоплазмы, окружающей спермин, основывалось на исследовании фиксированного материала на котором, как позже выяснилось, не удавалось сохранить нежную цитоплазму. Представление о спермиях-ядрах, а не клетках долго держалось в науке, так как оно укрепляло распространенную морганистскую теорию о монополии ядра в передаче наследственных свойств. И только недавно работами советских ученых Финна, Кострюковой и др. на живом, нефиксированном материале было доказано наличие хорошо сформированных спермиев — клеток. Работы Кострюковой и Черноярова показали также что спермин, несмотря на отсутствие специальных органоидов, обладают способностью активного передвижения.
Вопреки существовавшему раньше убеждению в пассивном переносе спермиев токами цитоплазмы в пыльцевой трубке, исследование живых объектов показало, что спермин активно передвигаются по направлению роста пыльцевой трубки (к яйцеклетке) и притом против течения цитоплазмы в ней.
Взаимодействие половых клеток, а также направленный рост пыльцевых трубок были давно известны науке. Мичурин и мичуринцы внесли существенное дополнение в понимание этого явления учением об избирательности оплодотворения: было установлено, что из смеси пыльцевых зерен (своих и чужих) не все в равной мере подходящие для слияния с данной яйцеклеткой данного цветка; в зависимости от вида партнеров, от их возраста и пр. изменяется их взаимная избирательность, что сказывается на свойствах потомства. Избирательность оплодотворения свойственна в равной мере и животным организмам.
Процесс оплодотворения оказывает огромное влияние на дальнейшее развитие оплодотворенных клеток; из них формируются новые органы нового организма.
Иное состояние живого тела, иной его обмен веществ выражается при этом морфологически в формировании другого (двойного) числа хромосом при делении клеток развивающегося спорофита, как мы видели и раньше.
Процесс оплодотворения оказывает, однако, влияние не только на те клетки, которые участвуют в оплодотворении. Мичуринская наука впервые показала, что процесс оплодотворения нельзя сводить к слиянию двух клеток. В процессе оплодотворения — своеобразного взаимного обмена веществ — принимают участие не только те спермин, которые сливаются с яйцеклеткой; другие мужские элементы, другие пыльцевые трубки, которые считались «лишними», оказывается, участвуют в процессе взаимодействия с тканями цветка.
Одной из многочисленных сторон влияния оплодотворения не только на Половые клетки, но и на другие клетки тела является формирование плода из завязи.
Издавна известно, что в результате оплодотворения у покрытосеменных одновременно с развитием оплодотворенных клеток, ведущих к формированию семени, наступают резкие изменения и в близлежащих тканях, особенно в завязи. Изменения эти ведут к развитию из завязи плода, в котором заключены семена, так же, как в момент оплодотворения были заключены в завязи семяпочки.
В процессе эволюции плоды играют новую роль, наряду с защитой скрытого в них развивающегося зародыша будущего нового спорового поколения: плоды также часто способствуют распространению семян, расселению растительных организмов. В связи с этим у плодов, как и у семян, выработались весьма разнообразные приспособления. Приспособления к распространению плодов и семян связаны опять-таки с окружающей природой. При переносе семян и плодов принимают участие те же факторы, которые осуществляют перенос пыльцы.
Разнообразные волоски, пушки, крылатые выросты способствуют разнесению при помощи ветра как семян (тополь, ива, хлопчатник), так и плодов (одуванчик, клен, вяз) (рис. 14).

Многие мелкие семена (например, табака и др.) также подхватываются ветром.
Некоторые сухие плоды раскрываются дырочками или трещинами, из которых высыпаются семена, когда растение раскачивается на ветру (мак, дурман и др.) (рис. 15). Животные разносят плоды и семена, снабженные прицепками, крючочками или имеющие клейкую поверхность.

Сочные плоды привлекают животных (птиц, млекопитающих) своим вкусом и часто яркой окраской (рис. 16). Они обычно содержат семена, хорошо защищенные плотной кожурой или частью околоплодника (косточки), предохраняющей их от повреждений при прохождении через пищеварительный тракт животного. Поедая плод, животное, таким образом, способствует распространению семян.

Итак, покрытосеменные, или цветковые, растения являются наиболее высокоорганизованными представителями листостебельных. По сравнению с голосеменными они приобрели в процессе эволюции ряд черт организации, повышающих общий уровень их жизнеспособности, их приспособительные возможности. К таким особенностям относятся:
1) наличие цветка с разнообразными приспособлениями к перекрестному опылению;
2) пестик, охраняющий семяпочки;
3) двойное оплодотворение;
4) плод, охраняющий семя и способствующий его распространению.
В подтипе покрытосеменных редукция гаметофита пошла еще дальше, чем у голосеменных.
Как сказано выше, она привела в типе семенных к потере видимого чередования поколений — половое поколение превратилось в часть спорофита, и потребовалось немало усилий коллективной мысли, чтобы выяснить гомологию органов покрытосеменных и споровых растений. Мы уже видели, что в это большое обобщение крупный вклад внесли отечественные ученые.
В подтипе покрытосеменных имеется два класса — однодольные и двудольные. Название это основано на различиях в строении зародыша семени: у однодольных растений зародыш содержит, кроме почечки (зародышевого стебелька с зародышевыми листочками) и корешка, один видоизмененный листочек — семядолю. Последняя прилегает к эндосперму и во время прорастания семени высасывает из него питательный материал. Единственная семядоля однодольных злаков называется щитком.
У двудольных растений в зародыше, кроме эмбриональной почечки и корешка, имеется два видоизмененных листочка. Эти два видоизмененных листочка — семядоли — часто накопляют в себе запас питательных веществ, как это бывает у бобовых и других.
Однодольные отличаются от двудольных и другими признаками: отсутствием камбия в сосудистоволокнистых пучках, иным расположением пучков в стебле (см. выше), иным расположением их также и в листе (в листьях двудольных сосудистоволокнистые пучки образуют сеть из ветвящихся «жилок», в листьях однодольных жилки идут более или менее параллельно друг другу). И однодольные, и двудольные возникли приблизительно одновременно, в начале мезозойской эры, и представляют собой две ветви развития, завоевавшие первенство среди других растений благодаря особенностям своей организации. Значение высших растений для человека, для всей жизни на Земле, как было указано во введении, чрезвычайно велико и многообразно.
Изучая особенности строения и жизнедеятельности растений и животных, мы все больше овладеваем искусством управлять живым миром, все больше подчиняем его интересам трудящегося человечества.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 