При изложении морфологии мы уже подчеркивали, что ретикулоэндотелиальная система — понятие не только или не столько морфологическое, сколько функциональное. Система эта, ограниченная морфологически (анатомически) ретикулярными и эндотелиальными клетками, многообразна по функциальной деятельности потому, что способна не только к восприятию различных веществ, вводимых извне или образующихся в самом организме в виде грубых взвесей или коллоидных растворов, но, главным образом, потому, что она активно участвует в процессах промежуточного обмена и разнообразных реактивных проявлениях организма. Сам принцип объединения рассеянных в различных тканях и органах клеточных элементов в одну систему по способности к поглощению и накоплению частиц разнообразных веществ говорит только о преимущественном значении этой функции ретикулоэндотелиальной системы как наиболее очевидного и изученного показателя деятельности всей системы. Эта функция граничит и не может быть полностью отделена от функции фагоцитоза, с которой непосредственно связаны многие другие проявления сложной функциональной деятельности ретикулоэндотелия.
Иллюстрацией к сказанному являются экспериментальные наблюдения при парентеральном введении крови различным животным. В этом отношении не потеряли и не потеряют своего значения классические опыты И. И. Мечникова с введением в брюшную полость морской свинки дефибринированной гусиной крови. И. И. Мечников с полной очевидностью доказал, что эритроциты, введенные под кожу или в брюшную полость животных, не изменяются в перитонеальной жидкости. Но уже в течение ближайших часов амебоидные клетки захватывают большую часть эритроцитов, которые подвергаются влиянию содержимого таких фагоцитов. Описания наблюдавшихся в этих опытах процессов фаголиза и фагозитоза так последовательны, так тонко характеризуют функциональную деятельность макрофагов и морфологические изменения при этом, что все последующие исследования и описания являются только частными дополнениями к ним. Заслуживает внимания тот факт, что через несколько дней после впрыскивания (3—4 дня) брюшная лимфа заключает одни вновь прибывшие лейкоциты, не содержащие ни красных шариков, ни их остатков. Чтобы отыскать макрофаги, поглотившие красные шарики, приходится вскрыть морскую свинку, и тогда их в большом количестве находят в железистой части сальника, в мезентериальных железах, в печени и селезенке. Они довольно легко узнаются, благодаря характерному виду остатков красных шариков. Поглотив их, макрофаги переходят из перитонеальной жидкости в вышеуказанные органы, где и доканчивают пищеварение. В печени они имеют вид больших одноядерных клеток, часто с очень сильно развитыми отростками. В таком состоянии они вполне напоминают звездчатые клетки Купфера.
И. И. Мечников говорил: «Избранный нами примеру разрушения гусиных красных шариков макрофагами морской свинки может служить прототипом для резорбирования организованных элементов вообще» К Наблюдениями Нейсера и опытами В. К. Высоковича было доказано, что микробы, попавшие в кровь, вскоре могут быть обнаружены в эндотелиальных клетках, главным образом, в печени. Бактерии, поглощенные клетками ретикулоэндотелиальной системы, как и эритроциты чужеродной крови, подвергаются постепенному разрушению. Ретикулоэндотелий других органов также принимает участие в фагоцитозе. Ряд исследователей (Розенталь — Rosenthal, Высокович, Кузуноку — Kusunoku и др.) показал, что основная масса бактерий поглощается эндотелием печени, в несколько меньшей степени селезенки, еде менее ретикулоэндотелием костного мозга, затем легких. То же самое можно сказать о фагоцитозе введенных в кровь животного чужеродных эритроцитов (Биллинг — Bieling и Изаак — Isaak, Герлах и др.). Первостепенное значение селезенки в процессе фагоцитоза микроорганизмов легко доказывается тем, что после спленэктомии поглощение последних резко замедляется.
И. И. Мечниковым при изучении процессов фагоцитоза были отмeчены два обстоятельства. Первое обстоятельство это то, что неподвижные амебоидные клетки, как крупные клетки селезеночной мякоти, лимфатических узлов и сальника, играют значительную роль в резервировании клеток.
По сути этим определяется единая функциональная система макрофагов, деятельность которых рассматривается И. И. Мечниковым с точки зрения «пищеварения в тканях», т. е. процессов обмена.
Изучению участия ретикулоэндотелиальной системы в белковом, липоидном, пигментном и других видах межуточного обмена посвящено очень большое количество работ. Однако направленность книги не позволяет даже коротко остановиться на этом чрезвычайно интересном, но обширном вопросе. Второе обстоятельство, отмеченное И. И. Мечниковым, заключается в том, что «после нескольких впрыскиваний крови морской свинки в брюшную полость золотой рыбки перитонеальная жидкость последней приобретает новые свойства. Если взять через 15 дней после первого вспрыскивания немного перитонеального экссудата золотой рыбки с каплей всплывшего на поверхность его серума и прибавить к этой жидкости красные шарики морских свинок, то они немедленно и очень сильно агглютинируются и вскоре растворяются...» И далее: «так как растворяющая или гемолитическая способность серума получается только в результате переваривания красных шариков внутри лейкоцитов, то становится вероятным, что растворяющее вещество есть не что иное, как внутриклеточный фермент, происходящий из лейкоцитов. Вопрос, к которому мы приступили, имеет существенное значение в исследовании рассасывания и зависящих от него явлений иммунитета». Таким образом, И. И. Мечников ставил явления иммунитета в зависимость от функции макрофагов, т. е. ретикулоэндотелиальной системы.
Остановимся на современном состоянии учения о значении ретикулоэндотелиальной системы в процессах выработки иммунных тел и месте их образования. Исследованиями Мурата (Murata), Биллинга и Изаака, вызывавших блокаду ретикулоэндотелия введением животным взвеси туши, коллоидного серебра или железа, было ясно установлено, что при блокаде и последовательном впрыскивании (введении) чужеродных эритроцитов вырабатывалось значительно меньше гемолизинов, чем у контрольных животных. То же доказали Биллинг и Изаак, Нейфельд и Мейер (Meyer), Зингер (Singer) и Адлер (Adler) по отношению к брюшнотифозным палочкам и пневмококкам. Ряд других исследователей установил явное ослабление образования бактериолизинов и агглютининов после блокады коллоидными красками. Ослабление еще резче выступает при спленэктомии и последующей блокаде. Однако далеко не всеми исследователями достигались близкие результаты, и Розенталь и Фишер (Fischer), Штанденат, Розенталь, Мозель и Петцаль, Алымов и Гуревич отрицают участие ретикулоэндотелиальной системы в выработке иммунных тел. Варшатова и Леонтьева также приходят к выводу, что блокада ретикулоэндотелия не влияет на образование антител, но объясняют это обстоятельство несовершенством методов блокады. Противоречия в выводах различных исследователей во многом разъясняются опытами Юнжблюта (Jungeblut) и Кобаяши (Kobajashi). Юнжблют показал, что иммунные животные после впрыскиваний туши нейтрализуют значительно меньшие количества токсина, т. е. блокада препятствует выработке иммунитета. Однако такое препятствие выражено только при первой инъекции антигена. При последующих инъекциях антигена влияние предварительного блокирования постепенно стушевывается и процессы выработки антител протекают как у контрольных (неблокированных) животных. Задерживающее влияние блокады заметно только, примерно, в течение первых суток и непродолжительность его объясняется (Н. Н. Аничков) быстро наступающей регенерацией клеток. Еще более четко показана кратковременность влияния блокады на выработку активного иммунитета в опытах Кобаяши. Одним из важных доказательств образования антител клетками ретикулоэндотелиальной системы являются опыты Кареля (Carrel), который нашел, что эритроциты козы, прибавленные к искусственной культуре костного мозга или лимфатических узлов морской свинки, вызывают образование в культуре гемолизина. Пршигоде (Przigode) наблюдал образование преципитинов и агглютининов в культуре селезеночной ткани.
Резюмируя данные относительно влияния ретикулоэндотелиальной системы на выработку иммунных тел, Н. Н. Аничков пишет, что «необходимо признать существование такого влияния, имеющего большое значение в смысле уничтожения патогенных микроорганизмов. Уничтожение микроорганизмов происходит частью внутри клеток ретикулоэндотелиальной системы, частью вне их при помощи антител, в выработке которых также принимает участие ретикулоэндотелий».
Явления иммунологической реактивности организма пересматриваются в настоящее время с точки зрения ведущего значения центральной нервной системы в регуляции всех процессов, происходящих в организме (А. Д. Адо, А. И. Гордиенко, П. Ф. Здродовский, С. И. Метальников, А. Д. Сперанский и др.). Наряду с тем, что ряд исследований показал определенное участие рефлекторного механизма в выработке антител, все же отмеченная уже И. И. Мечниковым роль ретикулоэндотелиальной системы в этом процессе не может умаляться. В этом смысле высказывается и П. Ф. Здродовский, который пишет: «Можно, однако, предполагать, что нервнорефлекторная продукция противотел должна иметь место, главным образом, при воздействии антигена на организм, уже находящийся в состоянии высокой иммунологической реактивности (отсюда эффективность малых доз антигена при ревакцинации). Напротив, при первоначальной перестройке реактивности организма под влиянием «первичной» иммунизации преимущественное значение, вероятно, имеют процессы ассимиляции антигена элементами тканей, продуцирующих противотела (отсюда необходимость массивных доз антигена при первичной иммунизации)». В таком же смысле высказывается и А. Д. Адо, который пишет: «Иными словами, рефлекторное антигенное раздражение может только регулировать уже наличный процесс выработки нормальных и, по-видимому, так же иммунных антител, но не в состоянии изменить интенсивность и качество работы аппарата, вырабатывающего антитела, в такой степени, как это происходит под влиянием обычной активной иммунизации. Думается, что для осуществления последней задачи, все же необходимы контакт и взаимодействие антигена с тканями, непосредственно вырабатывающими антитела и с тем сырым материалом (глобулинами крови), из которых эти антитела приготовляются».
После краткого обсуждения роли ретикулоэндотелиальной системы в развитии иммунитета, остается рассмотреть еще одну ее важную функцию в процессе кроветворения.
В настоящее время выработалось единодушное мнение о том, что клеточные элементы крови являются производными мезенхимы и спорные вопросы касаются, главным образом, последующих стадий развития. Однако при ближайшем рассмотрении становится ясным, что единодушие достигнуто только в понимании развития клеток крови из элементов мезенхимы в эмбриональном периоде. Для внеутробного периода решение этого вопроса оказалось более сложным. Стоит только пересмотреть схемы кроветворения, предложенные различными авторами, чтобы убедиться в чрезвычайном разнобое, который царит в представлениях об основном клеточном элементе, являющемся родоначальным в развитии клеток крови в постэмбриональном периоде,
Ряд исследователей вместе с Негели полагает, что таким родоначальным элементом является мезенхимальная клетка, которая выявляет свои потенции к кроветворению только в условиях патологии. В условиях физиологии кроветворение совершается только из существующих уже незрелых кровяных элементов — миэлобластов и лимфобластов.
А. А. Максимов полагал, что основой кроветворения являются элементы недифференцированного ретикулярного синтиция. Шиллинг с точки зрения триадиетической теории основным элементом кроветворения считает мезенхимальную клетку, производная от которой ретикулоэндотелиальная клетка является родоначальной для моноцитов. Фрейфельд также полагает основным элементом Мезенхимальную клетку, являющуюся родоначальной для ретикулоэндотелия. Из последнего развиваются уже материнские клетки элементов крови — эритрогонии, миэлобласты, лимфобласты, монобласты, плазмобласты и мегакариобласты. А. Н. Крюков производит клетки крови из гистиогенного элемента, а его ученики И. А. Кассирский и Г. А. Алексеев из ретикулоэндотелиальной клетки костного мозга. В последней схеме кроветворения И. А. Кассирский и Г. А. Алексеев называют исходный элемент гемогистобластом, а в последней работе (1952 г.) Г. А. Алексеев высказывает мнение о том, что лимфоидно-ретикулярная клетка является таким гемогистобластом. Наконец, в схеме, опубликованной в 1953 г., X. X. Владос говорит о ретикулярной клетке (ретикулярном синтиции) стромы кроветворных органов, как об основной и о гемогистиобласте, как о трансформированной ретикулярной клетке и т. д.
Несмотря на значительные различия, во всех мнениях можно найти то общее, что за основной элемент кроветворения принимают клетку тканевую, мезенхимальную или производную от нее.
Принимая во внимание это обстоятельство, казалось можно было бы игнорировать частные разногласия, если бы они ограничивались только терминологией и не выявляли бы более глубоких расхождений в понимании ретикулоэндотелиальной системы и ее функций. Для примера приведем мнение X. X. Владоса из последней его работы (1953 г.). «Для правильного понимания процесса кроветворения у человека во внеутробном периоде его жизни особое значение имеет выявление генетических взаимоотношений между недифференцированными элементами соединительной ткани, ретикулярными клетками и родоначальными кровяными клетками. Следует признать правильным мнение А. Максимова, что в гемопоэзе играет роль ретикулярный синцитий — строма кроветворных органов, а отнюдь не ретикулоэндотелий, как ошибочно указывают зарубежные авторы. В пользу этого положения говорят исследования гистологов (А. Максимов и его сотрудники, А. А. Заварзин и др.), доказывающие, что эндотелий, выстилающий синусы костного мозга, лимфатических узлов и селезенки, а стало быть и береговые клетки Зигмунда по своему гистогенезу принадлежат к ретикулярной ткани. Ретикулярные клетки гистогенетически связаны с мезенхимальными элементами эмбрионального периода и с соединительной тканью во внеутробной жизни. Лишь незначительная часть ретикулярных клеток, необходимая для удовлетворения нормальных потребностей организма, обладает в физиологических условиях гемопоэтической потенцией. Таким образом, воззрение А. Максимова, одно время ( в своих ранних работах) утверждавшего, что «мезенхимальные клетки» способны к кроветворению, так же как и представления дуалистов (Негели и др.) считающих «элементы мезенхимы» родоначальной кровяной клеткой, должны быть признаны несостоятельными: в физиологических условиях в период внеутробной жизни никаких элементов мезенхимы обнаружить не удается. Как было сказано, более правильным является взгляд, согласно которому во внеутробной жизни организма человека в кроветворении участвуют ретикулярные клетки».
В работе X. X. Владоса, многое, по крайней мере для нас, остается неясным. Нам кажется, что подавляющее большинство авторов под термином «мезенхимальные клетки», «элементы мезенхимы» имеют в виду не эмбриональную мезенхиму, а ее ближайшие производные во внеутробной жизни, и если элементов мезенхимы нет, как указывает X. X. Владос, в физиологических условиях, то их нет во внеутробной жизни вообще, ибо неоткуда им тогда взяться и в условиях патологии. Как понимать, что ретикулярные клетки в эмбриональном периоде гистогенетически связаны с мезенхимальными элементами, а во внеутробной — с соединительной тканью? Нам кажется несомненным, что ретикулярные клетки не связаны с соединительной тканью, а представляют собой вид соединительной ткани, гистогенетически являясь производным именно эмбриональной мезенхимы. Что же такое строма кроветворных органов — ретикулярный синтиций, — как не элемент ретикулоэндотелиальной системы?
Что касается «эндотелия» синусов, то ведь сам автор повторяет почти общепризнанное мнение, что эндотелий, выстилающий синусы костного мозга, лимфатических узлов и селезенки, в отличие от эндотелия сосудов, принадлежит к ретикулярной ткани, и не только генетически, как считает X. X. Владос, так как такое же происхождение имеет и эндотелий сосудов, но, главным образом, по своим функциональным возможностям. Следовательно, это только термин, которым подчеркивается, что такие ретикулярные клетки синусов являются в то же время покровными. Обладая, следовательно, одинаковыми функциями и клетки стромы, и клетки синусов относятся к ретикулоэндотелию и ретикулоэндотелиальной системе в узком смысле. Если полагать, что ретикулярные клетки стромы кроветворных органов обладают гемопоэтической потенцией, то нет оснований отрицать такую же возможность и ретикулярных клеток синусов. Вместе с тем, X. X. Владос говорит, что лишь незначительная часть ретикулярных клеток, необходимая для удовлетворения нормальных потребностей организма обладает в физиологических условиях гемопоэтической потенцией. Что же это за часть, которая отличается от другой части при тех же условиях? По-видимому, признавая правильность мнения А. Максимова, надо согласиться с ним до конца и говорить не о «ретикулярном синтиции», а о «недифференцированном ретикулярном синтиции», или еще лучше о тех ретикулярных клетках, обладающих потенцией к кроветворению, которые А. Максимов и его ученики называют элементами «индифферентной мезенхимы». Эти клетки расположены вокруг сосудов наряду с блуждающими клетками в покое, т. е. гистиоцитами и фиброцитами. Клиницистам и патогистологам хорошо известно, что не только при системных поражениях кроветворных органов, но и при различных инфекционных процессах, протекающих с гиперергической реакцией организма, могут возникать очаги кроветворения за пределами кроветворных органов. Откуда же, из каких элементов они возникают, если потенцией к кроветворению обладают по X. X. Владосу только элементы ретикулярного синтиция стромы кроветворных органов?
Противоречия в вопросах кроветворной функции ретикулоэндотелиальной системы далеко не исчерпываются приведенным примером, который в определенной мере свидетельствует уже о недостаточности наших представлений и знаний в этом важном разделе физиологии. Несмотря на это, многочисленные наблюдения позволяют нам высказать мнение, которое сложилось у нас при изучении физиологии и патологии кроветворения.
На протяжении всего изложения раздела о ретикулоэндотелиальной системе мы старались, с одной стороны, подчеркнуть разнообразие составляющих ее клеточных элементов и их общность, а с другой, показать, что элементы этой системы, которые мы обнаруживаем в тканях при гистологическом исследовании или в пунктатах из кроветворных органов, являются уже в большей или меньшей мере дифференцированными в морфологическом облике и особенно в функциональной направленности. Способность тех элементов, которые обозначаются как гистиоциты, развиваться только в определенном отношении в смысле возможности их превращения в более дифференцированные элементы типа фибробластов и фиброцитов признается подавляющим большинством исследователей. Мы имеем основания полагать, что не только эти элементы ретикулоэндотелиальной системы, но и те, которые обозначаются как ретикулярные клетки, в том числе и стромы кроветворных органов, являются уже элементами с определенной степенью дифференциации морфологической и функциональной. Являясь элементами опорной ткани кроветворных органов, они помимо этой роли выполняют другую, составляя существенную часть ретикулоэндотелиальной системы, как системы защиты организма, к которой в основном направлено разнообразие ее функций. На первый план при осуществлении этой роли выступают три процесса: гиперплазия ретикулярных элементов, усиление их фагоцитарной способности и выработка антител (иммунитет и аллергия). Эти три процесса взаимосвязаны, но под влиянием создающихся условий я состояния реактивности могут преобладать то усиление функциональных возможностей, то процессы размножения с тенденцией к отделению клеток в виде блуждающих макрофагов, то без такой тенденции в виде обратимой реактивной гиперплазии — реактивного ретикулоза, то в виде глубокой патологии е необратимой, нарастающей гиперплазией — острого или хронического ретикулоза или ретикулоэндотелиоза как системного поражения, с большим или меньшим участием упомянутого выше еще менее дифференцированного клеточного элемента «индифферентной мезенхимы». Никакой тенденции к кроветворению ретикулярные клетки не выявляют. В случаях изменения реактивности ретикулоэндотелиальной системы при гиперергических реакциях, когда может наблюдаться миэлоидное кроветворение в лимфатических узлах (А. И. Абрикосов, Е. Я. Герценберг и С. И. Ладыженская, Блюм и Земан — Seemann и др.), последнее совершается в зародышевых центрах, а не повсеместно в ретикулярном синтиции. Даже в выраженной патологии кроветворения при хроническом миэлозе, А. И. Абрикосов подчеркивает развитие миэлоидной ткани в синусах узлов, а не в ретикулярном синтиции.
Экспериментальные, клинические наблюдения и патогистологические исследования при различных условиях, в том числе и при системных заболеваниях кроветворных органов, позволяют присоединиться к мнению А. Максимова о существовании среди ретикулярных клеточных элементов еще менее дифференцированных, по крайней мере в функциональном отношении, клеток, обладающих полипотентностью в превращениях в другие клеточные элементы, которые он обозначал как элементы «индифферентной мезенхимы».
В процессе деления, в зависимости от условий среды, создается функциональная и морфологическая направленность развития этих элементов в сторону кровяных или юных ретикулярных клеток. При дифференциации этой «индифферентной мезенхимальной клетки» в кровяную под влиянием тех или иных причин она проходит промежуточную стадию, которая, обладая еще чертами этой индифферентной клетки, приобретает не только в функциональном, но и в морфологическом отношении особенности, свойственные развивающемуся из нее виду кровяного элемента — гемоцитобласту, проэритробласту, промегалобласту. Такие клетки в общем дифференцируются как гемогиетиобласты, под которыми одними авторами подразумеваются клетки Феррата, другими, как было указано выше, клетки ретикулярного синтиция кроветворных органов или лимфоидно-ретикулярные клетки. Гемогиетиобласты под влиянием условий среды (процессов обмена, анатомических отношений, особенности раздражителя и т. п.) получают направление развития в ту или другую кровяную лимфоидную предстадию. При физиологических условиях постэмбрионального кроветворения указанные промежуточные стадии встречаются очень редко. Для обычных потребностей организма, существующего в нормальных условиях, а часто и при значительно повышенной потребности в условиях патологии, выработка кровяных элементов достаточно- обеспечивается наличием уже более зрелых, но способных к делению миэлоидных и лимфоидных клеток в кроветворных органах — лимфобластов, лейкобластов, миэлоцитов и эритробластов. Под влиянием изменившихся условий, сущность которых не выяснена и представляется различно разными авторами, наряду с усилением пролиферации уже имеющихся в кроветворных органах клеток, происходит также производство и дифференциация клеток из элементов «индифферентной мезенхимы». Таким образом, принципиально различия в кроветворении при физиологических и патологических условиях касаются, главным образом, количества, а не качества. Соответственно нашим наблюдениям и воззрениям, процесс кроветворения может быть выражен следующей схемой (табл. II).
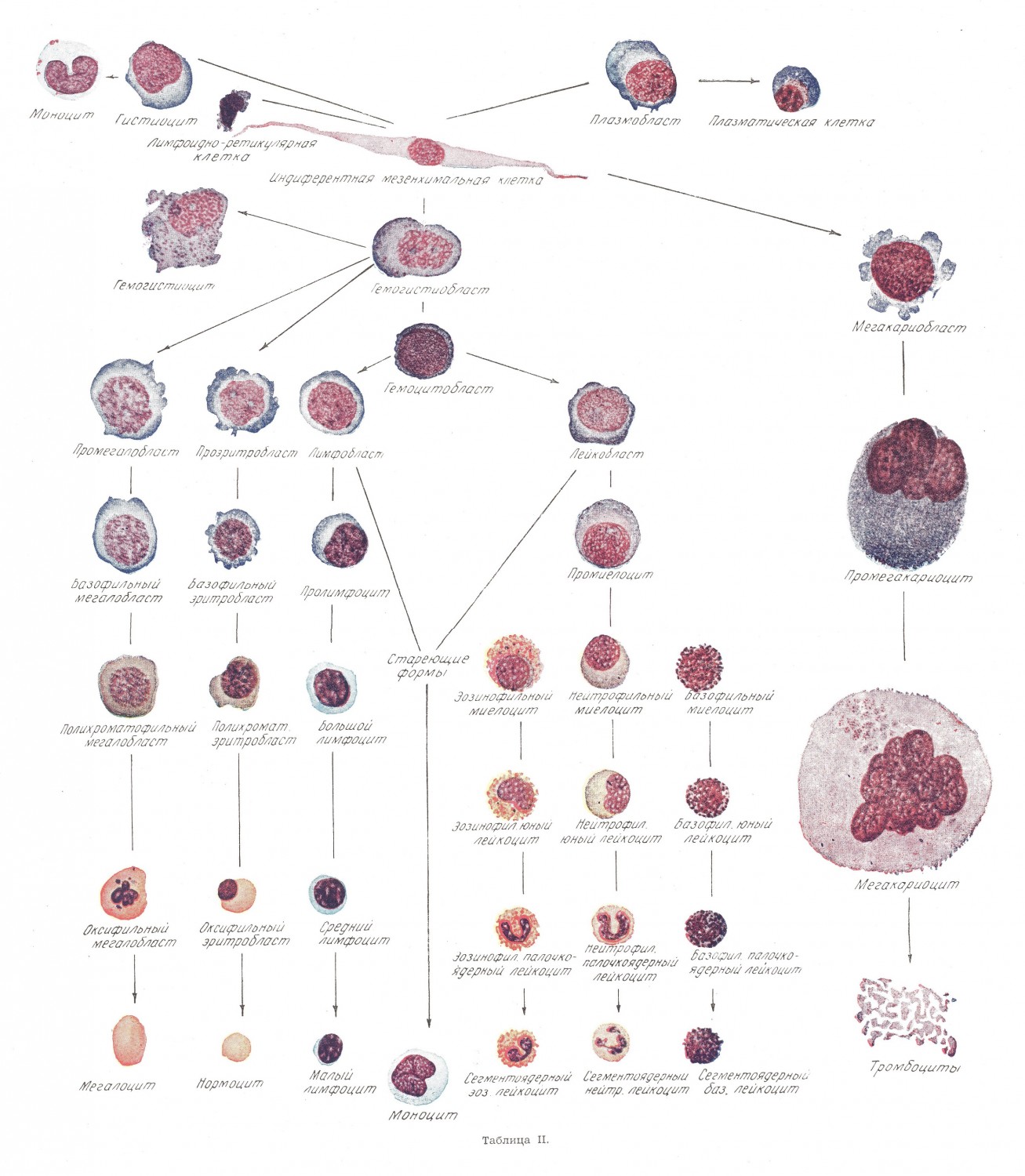
В этой схеме кверху от «индифферентной мезенхимальной клетки» расположены производные от нее тканевые элементы, редко проникающие в ток крови; ниже — кровяные элементы в последовательной их дифференциации. Существует большое количество различных схем кроветворения других авторов которые мы не обсуждаем, поскольку для рассматриваемых нами вопросов это не имеет сколько-нибудь существенного значения.
Какой бы схемы не придерживаться, нет разногласий (за редким исключением, например, схема Н. М. Николаева) в том, что в результате превращения и дифференциации тканевого элемента, входящего в состав ретикулоэндотелиальной системы, образуются зрелые кровяные элементы, обладающие хорошо очерчеными морфологическими особенностями и вполне определенной функцией. Специальные зрелые зернистые клетки — сегментоядерные нейтрофильные лейкоциты обладают выраженными свойствами фагоцитов. И. И. Мечников в отличие от фагоцитов — макрофагов ретикулоэндотелиальной ткани назвал их микрофагами. Он же описал различия в близкой по существу функциональной деятельности этих двух видов фагоцитов в следующих выражениях: «Таким образом, макрофаги преимущественно захватывают животные клетки, как кровяные шарики, семенные тела и всякие другие элементы животного происхождения. Между заразными микробами макрофаги предпочитают тех, которые вызывают хронические болезни, как проказу, туберкулез или актиномикоз, а также и тех, которые животного происхождения. В эту последнюю категорию входят амебоидные паразиты перемежающейся и техасской лихорадок и трипанозомы. Макрофаги могут также поглощать бактерии острых болезней, но за немногими исключениями, вмешательство их не имеет большого значения.
Микрофаги, наоборот, преимущественно действуют при острых заболеваниях. Они вовсе, или только незначительно включаются в борьбу с животными клетками. Так, они только изредка захватывают красные шарики того же или чуждого животного мира. Они также обнаруживают отрицательное отношение к паразитам животного происхождения и к некоторым бактериям, вызывающим хронические заболевания. В то время как макрофаги с большой жадностью схватывают лепрозных бацилл, микрофаги делают это только в виде исключения.
Морфологическим и физиологическим отличиям двух больших категорий подвижных фагоцитов (лейкоцитов) также соответствует различие состава их растворимых ферментов.
Кажется очень вероятным, что у одного и того же животного вида существуют два различных цитаза. Один из них действует гораздо сильнее на животные клетки, чем на бактерии, это — макроцитаз, находящийся в лимфатических органах и в серуме крови. Благодаря ему экстракт или настой селезенки, сальника и лимфатических желез более или менее легко растворяет красные шарики; но в то же время эти экстракты или настои не способны разрушать бактерий. Когда макрофаги схватывают красные шарики, снабженные ядрами, то они их вполне переваривают вместе с ядром, представляющим такое сильное сопротивление. Но когда те же фагоциты поглощают наиболее удобоваримых микробов, как холерные вибрионы, то действие их слабо. Вибрионы остаются довольно долго живыми, не превращаясь в зерна, и только с большим трудом перевариваются и разрушаются. Цитаз микрофагов, или микроцитаз, отличается обратными свойствами. Он легко переваривает многих микробов, но почти или вовсе не действует на красные шарики и другие животные клетки» К
В настоящее время известно, что течение инфекционных заболеваний — острое или хроническое — определяется не только свойствами возбудителя, но и состоянием реактивности организма. Следовательно, преимущественное участие в патологических реакциях макрофагов или микрофагов также находится в зависимости от реактивности организма. Таким образом, в организме существуют две категории клеточных элементов — тканевые ретикулоэндотелиальные и кровяные, приспособленные для борьбы с чрезвычайными раздражителями и сложившиеся в процессе эволюции в единую, мощную систему защиты организма.
Отсюда вытекает и отношение, в котором находится лейкоцитарная картина крови к тем патологическим процессам, которые наиболее часто и ярко связаны в приспособительных реакциях организма с включением описанной мощной защитной системы. Из всего изложенного вытекает, что при мобилизации средств защиты в случаях острого поражения организма, особенно при инфекционных заболеваниях, которые протекают с явлениями местного воспаления, совершенно очевидна большая потребность в лейкоцитах и большая их убыль. Будет ли это осуществляться с помощью специального фактора (В. Менкин), или с помощью других продуктов распада, но происходящая сначала перераспределительная реакция, зависимая от регулирующего влияния нервной системы, сочетается или сменяется повышением функциональной деятельности кроветворных органов, главным образом, костного мозга с лейкоцитозом и сдвигом в лейкоцитарной формуле крови.
В случаях так называемых хронических инфекционных заболеваний, т. е. заболеваний, протекающих в сенсибилизированном организме, особенно сопровождающихся так называемым специфическим воспалением, на первый план выступает тканевая реакция со стороны ретикулоэндотелиальной системы с небольшими сравнительно изменениями со стороны нейтрофильной картины крови, но иногда с качественными изменениями других клеточных элементов (эозинофилия, гистиоцитемия и др.).
Все сказанное выше позволяет нам подойти к освещению основного вопроса, поставленного в книге—каково же значение лейкоцитарной картины крови в клинике. Несмотря на то, что всеми признано огромное значение количественного и качественного лейкоцитарного состава крови для установления диагноза и оценки прогноза, в толковании биологической сущности лейкоцитарной реакции и ее отражения в картине крови нет общности. Основные положения, которыми широко пользуются и в настоящее время, исходят от одного из основоположников учения о картине крови — Шиллинга. Эти положения сформулированы им следующим образом: «...реакция лейкоцитов в случаях инфекции или интоксикации протекает закономерно по основным правилам, которые автор в соответствии с основными биологическими законами Арндт-Шульца для лейкоцитов формулировал следующим образом:
1. Незначительные раздражения вызывают лишь функциональные изменения лейкоцитарной картины, средние раздражения являются формативными, действуя через лейкопоэтические органы; сильные влияют также на образование отдельной клетки; самые сильные раздражения действуют угнетающе, ослабляя центральные и разрушая периферические клетки.
2. В большинстве инфекционных процессов первыми реагируют на раздражение нейтрофилы, затем моноциты и в последнюю очередь лимфоциты: различие инфекционных картин крови зависит от взаимного сдвига (во времени) этих трех фаз и от колебания степени реакции отдельных групп, а также от появления, наряду с ними, редких форменных элементов».
В свете современных представлений о реактивности организма и течении патологических процессов в зависимости от последней, которые мы коротко обсудили ранее, приведенные положения Шиллинга не выдерживают критики. Значение «биологической кривой Шиллинга», отражающей в общем течение процессов взаимодействия микро- и макроорганизма при острых инфекционных, особенно септических заболеваниях сопровождающихся адэкватной реакцией костного мозга и регенеративным сдвигом лейкоцитарного состава крови, хотя и сохраняется, но трактовка ее Шиллингом не может быть принята. В руководстве по клинической гематологии мы приводили в качестве примера лейкоцитарную кривую при крупозной пневмонии у больного В., 30 лет, который заболел 13/Ѵ 1948 г. и 16/Ѵ был помещен в клинику (рис. 25).
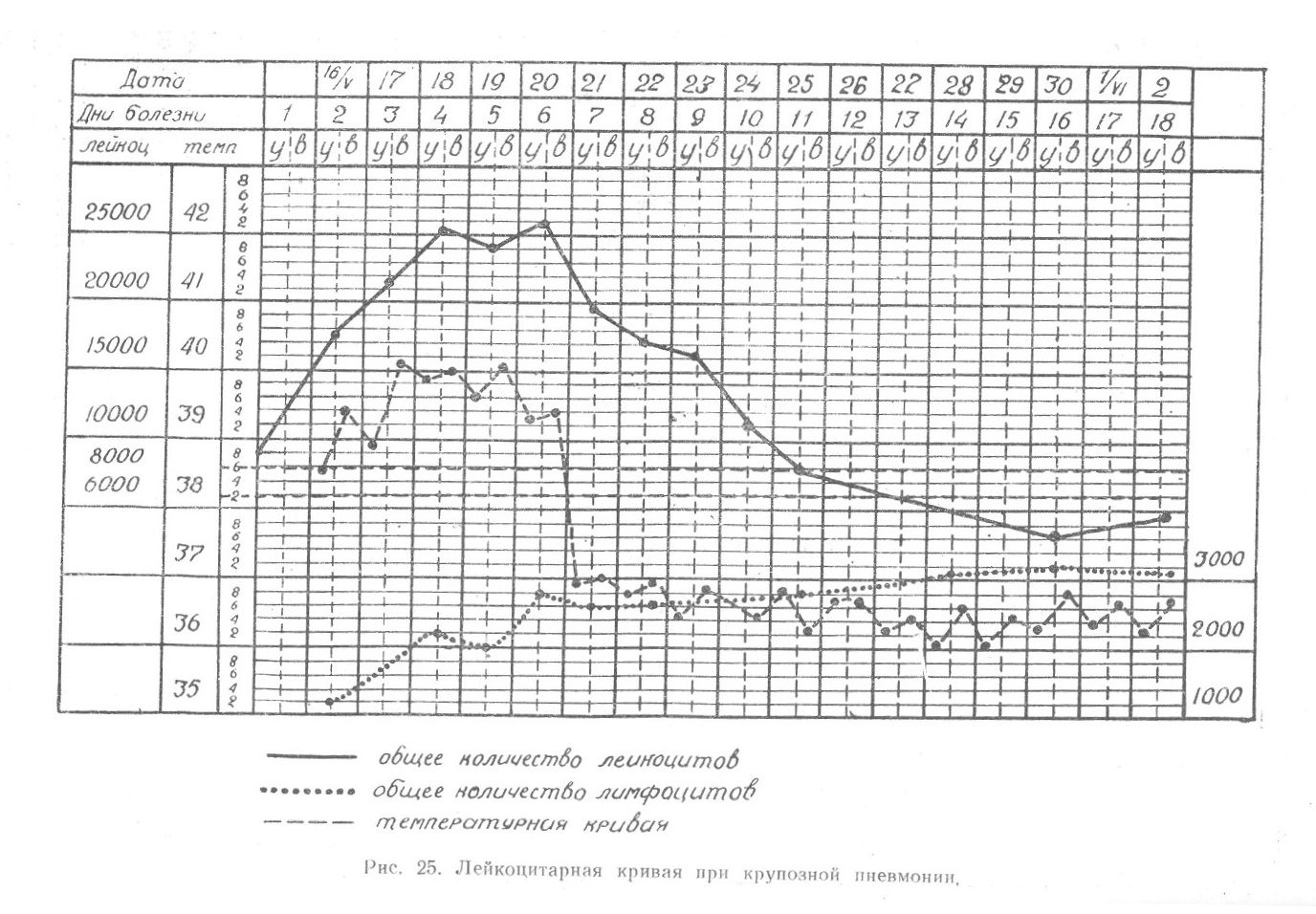
Изменения состава крови в течение крупозной пневмонии могут служить примером течения «биологической кривой» и при других инфекционных заболеваниях. Из приведенной кривой с очевидностью следует, что строгого чередования трех защитных фаз в борьбе организма с инфекцией, отражающихся на составе крови, в действительности вопреки мнению Шиллинга не существует. Впечатление о чередовании трех фаз может сседаться при учете только относительных чисел, т. е. процентного содержания отдельных форм лейкоцитов. В действительности же процент лимфоцитов и моноцитов в определенном объеме повышается только потому, что снижается, приближаясь к норме, процент нейтрофилов, за счет которых падает общее количество лейкоцитов. При переводе на абсолютные числа будет ясно, что количество нейтрофилов падает, aj число других форм лейкоцитов остается почти неизменным. В некоторых; случаях происходит увеличение и абсолютных чисел лимфоцитов и моноцитов, возможно и за счет усиления функциональной деятельности лимфатических узлов, и за счет перераспределения.
Близки к взглядам В. Шиллинга и положения об изменениях состава нейтрофильных лейкоцитоз Г. И. Когана и Н. Н. Боброва, но мы все же останавливаемся на них, главным образом, в связи с примененной авторами терминологией. Они пишут: «Незначительная нейтрофилия при легком сдвиге влево указывает на легкую инфекцию, значительная при большем ядерном сдвиге — на тяжелую. Если одновременно с незначительной нейтрофилией имеется небольшой лейкоцитоз, это свидетельствует о легкой инфекции и вполне хорошей сопротивляемости кроветворного аппарата. Значительная нейтрофилия с резко выраженным гиперлейкоцитозом говорит о тяжелой инфекции и хорошей сопротивляемости, резкая нейтрофилия с небольшим лейкоцитозом — о тяжелой инфекции и ослабленной сопротивляемости кроветворного аппарата; тяжелая нейтрофилия при отсутствии лейкоцитоза — показатель тяжелой инфекции и отсутствия сопротивляемости организма. При оценке нейтрофилии необходимо учитывать ее качественный состав: чем больше молодых форм, тем вирулентнее инфекция и тяжелее прогноз; чем меньше среди нейтрофилов юных и палочкоядерных и чем больше сегментированных форм, тем легче инфекция и благоприятнее предсказание».
Вряд ли кто-либо усомнится в том, что лейкоцитарная реакция с более или менее выраженным изменением лейкоцитарного состава крови является результатом взаимодействия «чрезвычайного раздражителя» с организмом. Как же можно в таком случае считать, что эти изменения характеризуют только «легкую и тяжелую» инфекцию, т. е. ее вирулентность?
Тяжесть инфекции, если под этим понимать инфекционное заболевание, характеризуется не лейкоцитарным составом крови, а общим состоянием больного, степенью интоксикации с поражением и функциональным нарушением ряда органов и систем и в первую очередь центральной нервной системы. Нам кажется чрезвычайно неудачным термин «сопротивляемость кроветворного аппарата», — хорошая или плохая, если его употреблять даже по аналогии с термином «сопротивляемости организма» инфекции. Под последним подразумевается устойчивость организма к инфекции, но назначение кроветворного аппарата не сопротивляться инфекции, а реагировать на нее определенной реакцией, защитной для организма. Следовательно, речь может идти не о хорошей или плохой сопротивляемости кроветворного аппарата, а о более или менее выраженной его реакции. Еще менее оправданным кажется нам такая формулировка — «тяжелая нейтрофилия при отсутствии лейкоцитоза — показатель тяжелой инфекции и отсутствия сопротивляемости организма», если даже понимать отсутствие «сопротивляемости», как ареактивность, анергию. Авторы приводят следующий пример (табл. 22).
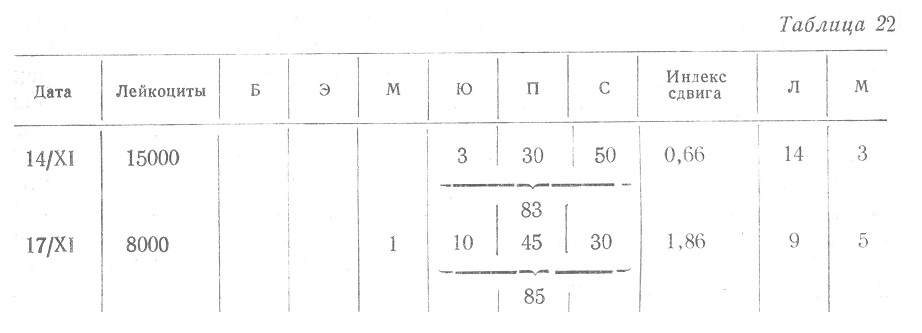
Можно ли сказать на основании этого примера, что 17/ХІ инфекция стала тяжелее, и прекратилась «сопротивляемость» организма и костного мозга? С большим правом можно думать, что сопротивляемость, т. е. реакция кроветворного органа не прекратилась и усиление сдвига влево по характеру не соответствует пассивному поступлению в кровь незрелых форм из косоного мозга. Можно было бы подумать только о том, что убыль лейкоцитов происходит интенсивнее, чем их восстановление за тот же промежуток времени, что потребность в лейкоцитах больше, чем может произвести при данных обстоятельствах кроветворный орган даже при высокой его активности. Один лейкоцитарный состав крови, без тщательной оценки клинической картины, примера которой авторы не приводят, не дает права делать столь ответственный вывод о прекращении сопротивляемости, о внезапном развитии анергии, о прекращении, или только резком ослаблении функциональной деятельности костного мозга. Этот пример является к тому же свидетельством невозможности учета лейкоцитарной картины крови для оценки прогноза, и особенно состояния реактивности организма, вне связи со всей клинической симптоматологией заболевания. В то же время в этом примере авторы незаметно для себя выходят за рамки ограниченного ими толкования гемограммы в пределах оценки функционального состояния кроветворного органа и распространяют толкование ее в смысле оценки сопротивляемости, т. е. реактивности организма. Подтверждением сказанного может служить пример, приведенный в руководстве И. А. Кассирского и Г. А. Алексеева.
«Больной Ч-ов, 51 года. Диагноз: прободной аппендицит. Операция. Констатирован гангренозный отросток. Брыжейка тонкой кишки и петли покрыты фибринозным налетом. В операционном ложе оставлен тампон. На следующий день после операции состояние больного признано тяжелым, угрожающим. Пульс 100 в минуту, язык сухой, живот очень вздут, газы не отходят. Температура 38,5. На третий день в состоянии больного трудно ориентироваться: живот вздут, отрыжка, но язык хороший, влажный; газы не отходят, но перистальтика имеется; небольшая желтушность:
Несмотря на то, что объективный status вызывает серьезные подозрения на развертывающийся перитонит, которого все ожидали на основании обнаруженного в брюшной полости распространенного воспалительного процесса, картина крови была исключительно благоприятной. Она казалась парадоксальной, совершенно не соответствующей состоянию больного: лейкоцитов — 7000, юных — 2%, палочкоядерных — 27%, сегментоядерных — 60%, лимфоцитов — 5%, моноцитов — 5%, эозинофилов — 1%. Токсогенная зернистость нейтрофилов +. Тельца Деле +. До операции: лейкоцитов — 20 100, юных — 2%, палочкоядерных — 25%, сегментоядерных — 60%, лимфоцитов — 8%, моноцитов — 5%.
Само собой понятно, что падение числа лейкоцитов нисколько не говорило за хороший прогноз, а скорее могло указывать на худшую форму перитонита, протекавшую с анергической реакцией кроветворения. Однако благоприятным симптомом в лейкограмме было то, что не появились миэлоцитарные элементы, главное же, что дегенеративная зернистость нейтрофилов была едва выражена (+). Мы рассуждали так: если это тяжелый перитонит, который протекает с анергической реакцией организма, то дегенеративная зернистость нейтрофилов в подобных случаях бывает особенно интенсивна [гнойные перитониты дают, как правило, резчайшую дегенеративную зернистость (++++) нейтрофилов и притом очень рано.
На основании трактовки лейкограммы больного мы склонились к благоприятному прогнозу. И действительно, при осмотре раны хирурги вскоре же подтвердили полное ограничение процесса (отсутствие каких-либо указаний на разлитой перитонит), и больной поправился».
Из этого, заслуживающего внимания примера, совершенно ясно, что ни об анергии кроветворения, ни об анергическом состоянии организма говорить не следует, поскольку были все явления распространенного воспаления — реакции, отнюдь не характеризующей анергию. Приведенные формулировки и примеры возбуждают важный вопрос — отображает ли лейкоцитарный состав крови реактивность организма и могут ли служить изменения состава крови показателем изменения его реактивности. Уже упоминалось о том, что лейкоцитарная картина крови слагается в результате процессов дегенерации в органах и тканях и регенерации в кроветворных органах. Совершенно ясно, что при всех процессах в организме, которые совершаются с помощью его защитной системы, принимают большее или меньшее участие лейкоциты. Последние в борьбе с вредоносным раздражителем гибнут в большем или меньшем количестве, что является поводом к стимуляции лейкопоэза в костном мозгу, отдающем повышенное количество выработанных лейкоцитов в кровь. В крови при этом обнаруживается лейкоцитоз без ядерного сдвига либо с большей или меньшей степенью сдвига в лейкоцитарной формуле, о чем уже было сказано ранее.
Количественный и качественный состав крови в совокупности отображают в достаточной мере функциональную деятельность кроветворных органов.
Степень реакции кроветворных органов и, в частности, костного мозга, определяется, с одной стороны, исходной реактивностью организма, его свойством так или иначе ответить на раздражение с участием защитных механизмов, и, с другой, реактивностью самого кроветворного органа. Нейтрофильная лейкоцитарная реакция, как выражение сложившейся в процессе эволюции защитной реакции организма отображает, таким образом, и состояние нормальной реактивности костного мозга, функцией которого является производство этого рода лейкоцитов, и нормальную исходную реактивность организма, которому присуща ответная реакция на определенный раздражитель с участием защитной системы. В таком смысле лейкоцитарная реакция отображает исходную реактивность организма и может быть показателем степени участия во взаимодействии организма и раздражителя кроветворной части системы защиты. Диапазон колебаний количественных показателей лейкоцитарной реакции очень широк и при более или менее однородных патологических процессах известны лейкоцитозы до 100 000 лейкоцитов в 1 мм3 со сдвигом формулы влево, и, едва превышающие верхние пределы нормы, лейкоцитозы в 10000—11000 в 1 мм3 с такого же рода сдвигом формулы влево. Изложенное выше указывает, что всякий лейкоцитоз — умеренный или высокий отображает нормальную реактивность кроветворного органа. Степень лейкоцитоза и ядерного сдвига или иначе степень выраженности лейкоцитарной картины крови, отображает только интенсивность лейкоцитарной реакции в зависимости от характера раздражителя, условий, при которых производится раздражение, исходной реактивности и анатомического состояния кроветворного органа, который принимает участие в ответной реакции организма. Следовательно, степень лейкоцитоза является только количественным выражением нормальной реактивности костного мозга. Поскольку обычный воспалительный процесс, как было указано выше, представляет собой присущую нормальному организму сложную защитную реакцию, то, естественно, при его возникновении можно говорить об адэкватной ответной реакции организма на определенное раздражение, свидетельствующей о нормальной реактивности организма по отношению к данному раздражителю.
Нейтрофильный лейкоцитоз является одним из очевидных и ярких выражений такого состояния и можно, казалось бы, сделать вывод, что в таком случае нейтрофильный лейкоцитоз может служить показателем нормальной реактивности организма, как это обычно и считают. Отсюда естественно сделать вывод, что отклонения в лейкоцитарной реакции, в лейкоцитарном составе крови могут служить показателем изменения реактивности. Вместе с тем это далеко не так просто и, как об этом будет сказано далее, инфекция, послужившая поводом к развитию обычного воспаления, так же как и продукты распада в воспалительном очаге, приводят к возникновению нового иммунобиологического состояния организма — сенсибилизации, аллергии, т. е. иной реактивности. В то же время развившаяся уже ранее обычная воспалительная реакция не исчезает, лейкоцитарная реакция продолжается, но лейкоцитоз уже не отображает действительного положения вещей, не отображает изменившейся иммунологической реактивности и остается только показателем наличия сохранившегося гнойно-воспалительного местного процесса. Легко можно себе представить и другое положение. В сенсибилизированном организме, где реакции протекают по другому типу — с преимущественным ответом ретикулоэндотелиальной ткани, следовательно, без нейтрофильного лейкоцитоза, может присоединиться влияние нового, отличного по своим свойствам, «чрезвычайного раздражителя». Тогда на фоне имеющихся проявлений гиперергической реакции той или иной интенсивности может возникнуть обычное воспаление с той или иной степенью нейтрофильною лейкоцитоза, и последний и тогда не явится показателем действительной реактивности организма. Значительное сложнее выяснение значения лейкопенического состава крови.
Не касаясь перераспределительных лейкопений, можно сказать, что последние оцениваются многими авторами как выражение понижения реактивности организма — ареактивности, анергии. Особенно характерным для состояния анергии многие авторы считают значительный сдвиг формулы влево до миэлоцитов при отсутствии лейкоцитоза. Реактивность организма в состоянии нормобиоза не представляется фиксированной и неизменной. Реакции организма могут быть неодинаковыми не только в зависимости от способа и условий приложения раздражителя, но и от возраста и физиологического состояния организма, от условий окружающей среды (Ш. Д. Мошкозский). Примером арективности организма при состоянии физиологической депрессии является феномен спячки. «Состояние зимней спячки характеризуется изменениями преимущественно депрессивного характера со стороны кровеносной и кроветворной системы (снижение кровяного давления, уменьшение крови, снижение лейкоцитов и пр.), со стороны эндокринной системы (понижение функции щитовидной железы, коры надпочечников, половых желез и передней доли гипофиза с усилением функции задней доли гипофиза и поджелудочной железы) и со стороны нервной системы (глубокое торможение центральной нервной системы, снижение тонуса симпатической системы, выключение терморегуляции и тонизирование парасимпатической системы)».
Мы привели эту характеристику зимней спячки (по А. М. Эмме) для того, чтобы подчеркнуть, с одной стороны, склонность при этом к лейкопении, а с другой, депрессивное состояние многочисленных жизненно важных систем, что определяет состояние близкое к анабиозу при предельном низком уровне обмена, сопровождающееся временным «затуханием» всех жизненных функций организма (П. Ф. Здродовский). Лейкопения в таком случае устанавливается в соответствии, по-видимому, не только с депрессивным состоянием кроветворных органов, но и с отсутствием более высокой потребности в лейкоцитах при отсутствии реакций взаимодействия организма и раздражителя, свойственных нормальной реактивности. Следовательно, в таких случаях чрезвычайно глубокой депрессии, лейкопения действительно является частичным, косвенным выражением ареактивности, анергии. Примером состояния депрессии у человека может служить алиментарная дистрофия, так называемая «болезнь голодания», изученная в период военной блокады Ленинграда. Всеми авторами при этом отмечена лейкопения. Правда, степень ее довольно широко вариирует по данным разных авторов. Д. М. Штейнберг указывает среднее число — 4800 лейкоцитов в 1 мм3; С. И. Шерман — ниже 6000 у 61,5—68% дистрофиков; И. М. Флекель — 5600, В. А. Свечников — 5700, причем в его наблюдениях меньше 4000 лейкоцитов было только у 14% больных. Таким образом, можно отметить, что лейкопения была при этом умеренной, в большинстве случаев в низких пределах нормы, если принять для последней 5000 — 6000 лейкоцитов в 1 мм3.
Еще больший интерес представляют при алиментарной дистрофии изменения состава крови при различных заболеваниях. При острых нагноительных процессах у дистрофиков В. А. Свечниковым и Т. И. Новоселовой был обнаружен гиперлейкоцитоз у пяти из 13 больных, умеренный лейкоцитоз — 10 000—15 000 — был у двух, выраженный — 15 000—20 000 — у двух и высокий — более 23 000 — у одного больного. У двух больных лейкоцитоз держался на верхних пределах нормы — 7000—8400 — и только у шести он был ниже 6000. Достойно внимания то обстоятельство, что в лейкоцитарной формуле при повышенном количестве лейкоцитов наблюдался нейтрофилез с умеренным сдвигом формулы влево. В тех случаях, где нагноение не сопровождалось гиперлейкоцитозом, не наблюдался ни нейтрофилез, ни сдвиг формулы влево.
Физиология ретикулоэндотелиальной системы, часть вторая
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
|
