О регуляции кроветворения, часть первая
Исследования Гоффа показали, что при введении воздуха в желудочки мозга наступает нейтрофильный лейкоцитоз с ядерным сдвигом влево и лимфопенией, длящийся около 24 часов. Исследования Гинцберга (Ginzberg) и Гейльмейера подтвердили это положение и показали в то же время у больных с органическими поражениями центральной нервной системы значительное нарастание количества ретикулоцитов в ближайшие часы с последующим снижением до исходных чисел или несколько ниже, примерно, через 24 часа. При отсутствии органического поражения центральной нервной системы ретикулоцитоз при тех же условиях не наблюдался.
Аналогичные данные были получены С. Н. Астаховым и М. Г. Аврутисом. Следует отметить, что Гаварцени и Бергетти наблюдали ретикулоцитоз при введении воздуха в полость мозговых желудочков у кроликов. Более широко были проведены исследования при энцефалографии В. П. Безугловым, Я. М. Павловским, Т. Ф. Фесенко, В. В. Черниковым и В. К. Эрмишем, которые получили примерно такие же данные как и другие исследователи с максимальной реакцией через 3—5 часов и (возвращением к нормальным числам в период от 10-ти до 24-х часов. В противоположность Гейльмейеру, они придают меньшее значение характеру патологического процесса в центральной нервной системе и большее — проникновению воздуха в желудочки. В соответствии с исследованиями Бальдучи (Balduzzi) и Пизани (Pisani), В. П. Безуглов, Я.М. Павлонский и другие авторы полагают, что изменения состава крови обусловливаются раздражением «центральных вегетативных аппаратов» как введенным воздухом, так и нарастающим давлением спинномозговой жидкости после пункции. Наконец, эти авторы считают, что изменения состава крови теоретически должно происходить по двум путям — по пути перераспределения и по пути раздражения гемопоэтических органов. Я. И. Минц и E. М. Фукс при вентрикуло- и энцефалографии также обнаружили лейкоцитоз с максимумом через 3 часа, с возвращением к исходному числу через сутки. Эти авторы полагают, что изменения в крови связаны с повышением внутричерепного давления, обусловливающим раздражение «центральных вегетативных аппаратов, расположенных вокруг третьего и четвертого желудочков».
Подобные результаты получил А. Я. Губергриц, который полагает, «что эти изменения числа лейкоцитов не являются чисто распределительного порядка, а, очевидно, вызваны какими-то, по-видимому, нейрогуморальными влияниями на костный мозг, исходящими из нервных центров, расположенных в головном мозгу.»
Иная картина была обнаружена А. Я. Губергрицем при энцефалографии у больных бронхиальной астмой, причем он считает, что здесь в первую очередь реагирует на введение воздуха в желудочки легко возбудимый у них вагус, который вызывает начальную лейкопению, а позже при значительном уже раздражении вегетативных центров, наступает вторично лейкоцитоз костномозгового происхождения.
Большой интерес представляют исследования, проведенные Т. С. Истамановой. Она показала зависимость изменений количественного состава крови под влиянием раздражения при энцефалографии и люмбальной пункции от состояния внутричерепного давления. Оказалось, что при энцефалографии и люмбальной пункции у людей с повышенным давлением ликвора количество ретикулоцитов также возрастало, но число эритроцитов уже в течение первого часа снижалось на 500 000 — 1 300 000. Этот факт Т. С. Истаманова считает возможным объяснить только перераспределением крови. Увеличение количества ретикулоцитов при сильной боли во время прокола, без искусственного изменения давления, колебания ретикулоцитов и эритроцитов в зависимости от состояния давления ликвора до вмешательства, приводят автора к выводу, что эти изменения являются следствием сосудистой реакции. Перечисленные данные позволили ей высказать предположение, что изменения со стороны крови при энцефалографии являются, может быть, следствием раздражения сосудодвигательного центра, а не спинального центра кроветворения. Последующие опыты Т. С. Истамановой, преимуществом которых является наряду с исследованиями крови изучение непосредственно изменений костного мозга, не решили вопроса о существовании специального центра, регулирующего кроветворение, но результаты их все же заслуживают внимания.
В этих опытах кролику, после предварительной резекции ребра и гистологического исследования костного мозга, в большую цистерну вводилась мелкодисперсная взвесь карболена. У контрольного кролика производились те же вмешательства и окципитальная пункция и не вводился только карболен. У всех животных, не исключая контрольных, уже через 30 минут начиналось нарастание числа эритроцитов до 1 000 000 в мм3, лейкоцитов и особенно отчетливо ретикулоцитов, достигавшее максимума через 1—2 часа и в ряде случаев уже через 4 часа снижавшееся до исходных чисел. При длительном (в течение нескольких месяцев) наблюдении выяснилось, что количество эритроцитов, лейкоцитов и ретикулоцитов повышалось вновь; на высоте подъема кривой через 3—3,5 месяца снова исследовался костный мозг. По сравнению с первыми исследованиями была отмечена активация как эритропоэза, так и, особенно, лейкопоэза. Через 5 месяцев после введения карболена у двух кроликов развились параличи, кахексия, и быстро стало уменьшаться количество гемоглобина и эритроцитов. Оба кролика погибли. Грубых анатомических изменений (абсцесс, кровоизлияние) в центральной нервной системе не было обнаружено, но более детальным гистологическим исследованием автор не располагал.
Т. С. Истаманова приходит к выводу, что начальные изменения в крови являются результатом только усиленного поступления форменных элементов из костного мозга в кровь, а более поздние изменения и длительное нарастание всех кровяных элементов являются результатом активации миэлопоэза в костном мозгу. Однако автор допускает, что эти изменения связаны с развитием hydrocephalus internus или с хроническим воспалительным процессом.
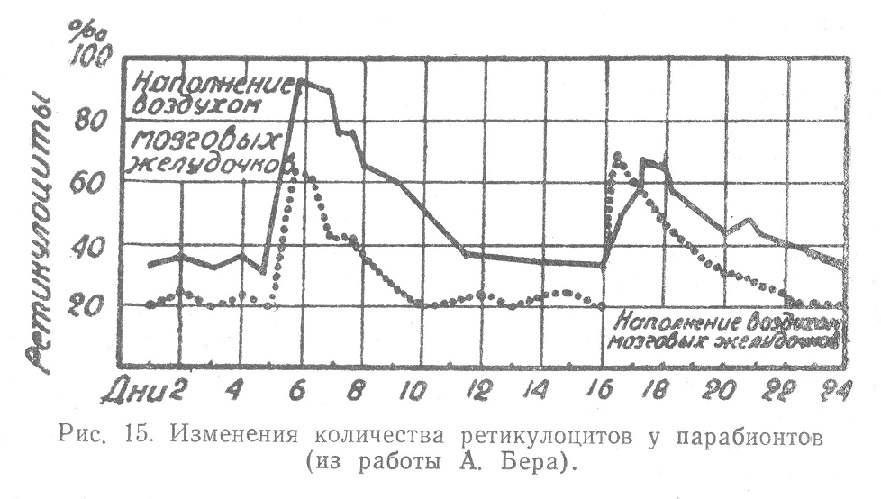
Наконец, большое значение для решения вопроса о влиянии центральной нервной системы на гемопоэз имеют опыты Бера (A. Beer) с парабиозом двух кроликов, при котором достигалось соединение двух полостей тела (соеlіо — анастомоз) с полным устранением нервных связей парабионтов. Исключив мышечное соединение, удалось добиться сращения по типу трансплантата, так что в рубце при очень скудно развитой капиллярной сети не мог происходить сколько-нибудь заметный обмен клеточными элементами крови. Вместе с тем обмен жидкостей сохранялся: при введении 1 мм3 10% раствора йодистого калия одному из парабионтов, уже через 40—60 минут можно было обнаружить йод в моче у партнера. Небольшое значение сосудов в парабиотическом мостике при этих гуморальных обменных отношениях доказывалось тем, что внутривенно введенный торотраст (6—8 мм3 на 1 кг веса тела) даже через 6—8 часов не обнаруживался рентгенологически у партнера, в то время как у животного, которому вводилась контрастная масса, получалось контрастное изображение печени и селезенки. Примерно через 4 недели после операции, когда устанавливался более или менее постоянный уровень кривой ретикулоцитов, одному из парабионтов путем окципитальной пункции наполняли воздухом мозговые желудочки. Уже через 3 часа наступало ясное нарастание количества ретикулоцитов, достигавшее в приводимом примере наибольшей величины через 9 часов (от 31 до 92%о). Через короткое время такой же подъем ретикулоцитов (от 20 до 64%о) наступал у парабионта, не подвергавшегося раздражению (рис. 15). В дальнейшем происходило медленное снижение количества ретикулоцитов до исходных чисел, которые устанавливались у партнера, не подвергавшегося раздражению, на несколько дней раньше, чем у подвергавшегося. Так как у первого и подъем ретикулоцитов был меньше, надо думать, что и раздражение у него было менее интенсивным. Опыт повторялся через некоторое время в обратном порядке с подобными же результатами. Последние могли быть истолкованы в том смысле, что в процессе нервного раздражения освобождаются вещества, под влиянием которых и происходят указанные изменения состава крови. Эти вещества поступают через парабиотический мостик от раздражаемого к не раздражаемому партнеру. Из этих опытов делается вывод, что в физиологических условиях при нервной регуляции крови между нервным раздражением и кровообразующими органами включаются вещества, действующие гуморально.
Мы привели ряд интересных экспериментальных и клинических исследований и наблюдений, которые должны были бы служить доказательством центральной вегетативно-нервной регуляции гемопоэза. Этими исследованиями с несомненностью показано влияние нервной системы на состав крови, но как только ими пытаются доказать такое влияние на кроветворение неизбежно выступают противоречия, неопределенность формулировки, а иногда и просто недооценка сущности процессов кроветворения. Хорошим примером может служить вывод В. П. Безуглова и его соавторов: «Среди путей, по которым центральные вегетативные аппараты изменяют состав крови, следует отметить влияние их на гемопоэтические органы, в наших случаях выразившиеся в том, что после вмешательства иногда появляются в периферическом русле крови клетки раздражения Тюрка и анизоцитоз, исчезающие обычно к 10—24 часам.» Следует заметить, что плазматические клетки (Тюрка), с какой бы точки зрения ни рассматривать их происхождение, не могут служить выражением усиления или угнетения процессов кроветворения. Их гиперплазия в кроветворных органах и некроветворных тканях и появление в крови происходит при определенных патологических условиях интоксикации или сенсибилизации.
В физиологических условиях их обычно в крови не находят, но изредка отдельные клетки проникают в кровь и количество их до 0,5% считается нормальным. Уже упоминалось о том, что количество этих клеток в крови, как правило, не отражает их количество в кроветворных органах и других тканях. В исследованиях указанных авторов единичные клетки Тюрка были обнаружены непостоянно только в двух из 18-ти случаев. Ни в какой мере не дает оснований для оценки состояния кроветворения и анизоцитоз, тем более учитывающийся на глаз. Но, если бы анизоцитоз и мог служить выражением состояния кроветворения, то и тогда авторы не имели бы права на какие-либо выводы, поскольку он был обнаружен только в одном из 18-ти случаев. Более достоверным признаком влияния на кроветворение мог бы служить ядерный сдвиг влево нейтрофилов, но незначительный и кратковременный сдвиг до палочкоядерных и при том далеко непостоянный, также не является доказательным. В лучшем случае он может быть объяснен выбрасыванием палочкоядерных нейтрофилов из синусов костного мозга как депо лейкоцитов. Такое мнение разделяет ряд авторов, в том числе В. Н. Черниговский и А. Я. Ярошевский, которые пишут: «Можно предполагать, что костный мозг сам является местом, где скапливаются и откуда выбрасываются лейкоциты.» То же можно повторить в отношении ретикулоцитов.
Одним из доказательств регуляции кроветворения нервной системой могло бы служить наличие нервных окончаний в паренхиме костного мозга. Следует отметить, что исследования иннервации костного мозга, по сравнению с такими же исследованиями других органов, явно недостаточны и неопределенны. Частично это объясняется недостатками методов, но в большей мере, по-видимому, анатомическим строением костного мозга. Недаром последний относится к «сосудистым органам». Характерно, что А. А. Заварзин и А. В. Румянцев подчеркивают очень простое строение костного мозга. Они пишут: «Основу его составляет сеть кровеносных сосудов, начинающихся артериями, которые проникают внутрь кости, переходят в капилляры, пронизывающие весь костный мозг и продолжаются в вены». При таком обилии кровеносных сосудов богато снабженных нервами, учитывая, что костный мозг заложен в полостях костей, разделенных костными балками, трудно решить, какую именно ткань иннервируют обнаруживаемые тончайшие нервные волоконца.
Среди немногочисленных исследователей иннервации костного мозга, только отдельные (де Кастро — de Castro, Оттоленги — Ottolenghi, Росси — Rossi) указывают, что нервные волоконца проникают непосредственно в паренхиму костного мозга. В. Н. Черниговский и А. Я. Ярошевский сообщают, что в самое последнее время нервные волокна в самой костномозговой ткани обнаружил Г. И. Чекулаев (1952), с работой которого нам познакомиться не удалось. Однако, даже если считать наличие нервных волокон в самой ткани костного мозга доказанным и согласиться с проблематичной схемой взаимоотношений между нервной, сосудистой системой и ретикулоэндотелиальной тканью, представленной в руководстве Е. Фрейфельд, то и тогда еще не станет вполне ясным, для каких целей предназначены эти нервные волокна и какую из многообразных функций ретикулярной ткани они регулируют. Наличие афферентной иннервации костного мозга признается доказанным исследованиями А. Я- Ярошевского. Не доказанным остается наличие функциональных нервов, т. е. то, что некоторые авторы полагают само собой разумеющимся,, поскольку не доказано прямое, непосредственное влияние нервной системы на кроветворение. Конечно, с таким положением может быть и не так легко согласиться, но об этом упрямо свидетельствуют экспериментальные и клинические факты. Возможно, что причины и объяснение их кроются в особенностях костного мозга как органа, основной функцией которого является постоянное размножение и созревание, т. е. производство организованных и сложных образований — разнообразных клеточных элементов. Последние только до известного предела остаются в синцитиальной связи, а затем отделяются и мигрируют в просвет венозных синусов и потому, по крайней мере в этом периоде, трудно полагать непосредственное влияние нервных волокон или их окончаний на отдельные клеточные элементы.
Доказав наличие афферентной иннервации внутренних органов, и в частности кроветворных, В. Н. Черниговский и А. Я. Ярошевский, естественно, поставили вопрос о роли и значении в регуляции системы крови рефлексов, возникающих при раздражении внутренних органов. Рефлекторному влиянию на состав крови уделялось и уделяется много внимания и по этому поводу имеются многочисленные исследования, на большей части которых мы не будем иметь возможности останавливаться. Изменений состава крови в связи с рефлекторными влияниями на кровеносные сосуды мы коснулись в предыдущей главе. Здесь считаем целесообразным остановиться более подробно на рефлекторных изменениях состава крови при раздражении рецепторов желудочно-кишечного тракта и так называемого, пищеварительного лейкоцитоза. В соответствующей главе мы подчеркивали, на основании собственных наблюдений и многочисленных экспериментальных данных, особенно опубликованных в последние годы, что существование так называемого пищеварительного лейкоцитоза, как физиологической реакции организма, не подлежит сомнению. Следует, однако, отметить, что эта реакция более выражена у животных, чем у человека, что полностью объясняется влиянием у последнего более совершенной центральной нервной системы. Естественно, что наряду с обилием работ, посвященных вопросам пищеварительной лейкоцитарной реакции, не было недостатка в гипотезах и теориях, созданных для объяснения причин и механизмов, ее обусловливающих.
Не оправдали себя предположения Вирхова (Virchow), Гоффмейсте-pa (Hoffmeister) и других авторов, о связи лейкоцитоза с усилением лимфопоэтической деятельности регионарных лимфатических и лимфоидных образований, как и предположения Левита (Lowit) и Габричевского о связи лейкоцитоза с явлениями химиотаксиса лейкоцитов. Не вполне разрешенным остался вопрос о связи пищеварительной лейкоцитарной реакции с деятельностью пищеварительных желез и особенно выделением соляной кислоты в желудке. По прямым или косвенным данным одних исследователей (Э. М. Гельштейн, В. Г. Чижиков, О. С. Часовников, Чиачио — Сіассіо, Панье — Pagniez и Рише — Pichet) связь пищеварительной лейкоцитарной реакции с выделением соляной кислоты в желудке как будто не оставляет сомнения, по данным других (Шабуневич — Szabuniewicz, Глазер) в возникновении лейкоцитоза выделение соляной кислоты никакого значения не имеет. Наблюдая два подъема на лейкоцитарной кривой при кормлении собак мясом, В. Г. Чижиков связывал первый подъем количества лейкоцитов с секрецией желудочного сока, второй — с всасыванием из кишечника продуктов переваривания мяса. Зависимость «пищеварительного лейкоцитоза» от выделения соков кишечных и поджелудочной железы наблюдали В. М. Соколов и Г. Н. Удинцев.
Уже В. М. Соколов отметил периодическое нарастание количества лейкоцитов также и при голодании и назвал его «периодическим лейкоцитозом». Последний он объяснял периодической деятельностью пищеварительных органов с отделением пищеварительных соков. Значительно позже Е. Мюллер и Петерсен доказали увеличение количества лейкоцитов при голодании в зависимости от периода больших сокращений желудка. Это обстоятельство они объясняли изменением сосудистого тонуса под влиянием раздражения того или иного отдела вегетативной нервной системы. На основании своих тщательных исследований, которые были подробно приведены ранее, А. Я. Губергриц пришел к выводу, что причиной различного направления лейкоцитарных реакций (в одних случаях с пищеварительным лейкоцитозом, в других с лейкопенией) на один и тот же пищевой раздражитель является различная установка в вегетативной системе у различных людей, а в разные дни у одного и того же индивидуума. Он полагает, что «действие приема молока следует рассматривать не только как воздействие его путем всасывания из кишечника в кровь, а как своего рода «удар» по вегетативной системе, которая вследствие различной своей «настройки» и обусловливает различное направление лейкоцитарной кривой». Однако понятия «вегетативная система», «вегетативная настройка и установка», «удар» по вегетативной системе являются настолько неопределенными, что ни в какой мере не объясняют ни сущности, ни механизма пищеварительных лейкоцитарных реакций, при изучении которых автор получил ряд интересных и важных данных.
Наконец, лейкоцитоз после приема пищи, в частности при «гемоклазическом кризе» Видаля, расценивался рядом исследователей как результат рефлекса со слизистой оболочки желудка (Н. Н. Иваненко-Чоповский, В. М. Шаверин, Е. Мюллер и Петерсен — Petersen, А. Я. Губергриц, А. Я. Ярошевский). Наиболее обширные и тщательные исследования рефлекторных влияний раздражения рецепторных полей на так называемую «систему крови» были предприняты в последние годы В. Н. Черниговским и его сотрудниками (А. Я. Ярошевский, Е. Л. Кан и С. П. Яковлев, О. С. Меркулова и О. И. Моисеева). А. Я. Ярошевский в эксперименте на кошках показал, что раздражение желудка, введенным через фистулу и раздутым при давлении 35—40 мм ртутного столба резиновым баллончиком, дает значительные колебания лейкоцитов. В первые 3—10 минут наблюдается увеличение, а через 30—60 минут снижение их количества ниже исходного уровня. Но уже ко второму, реже к третьему часу, наступает выраженный и стойкий лейкоцитоз, сохраняющийся в течение всего опыта, несмотря на то, что раздувание уже через два часа заканчивается. Наибольший прирост количества лейкоцитов составлял 91,5%, а наибольше снижение — 33,7%. Указанные изменения представлены на рис. 16.
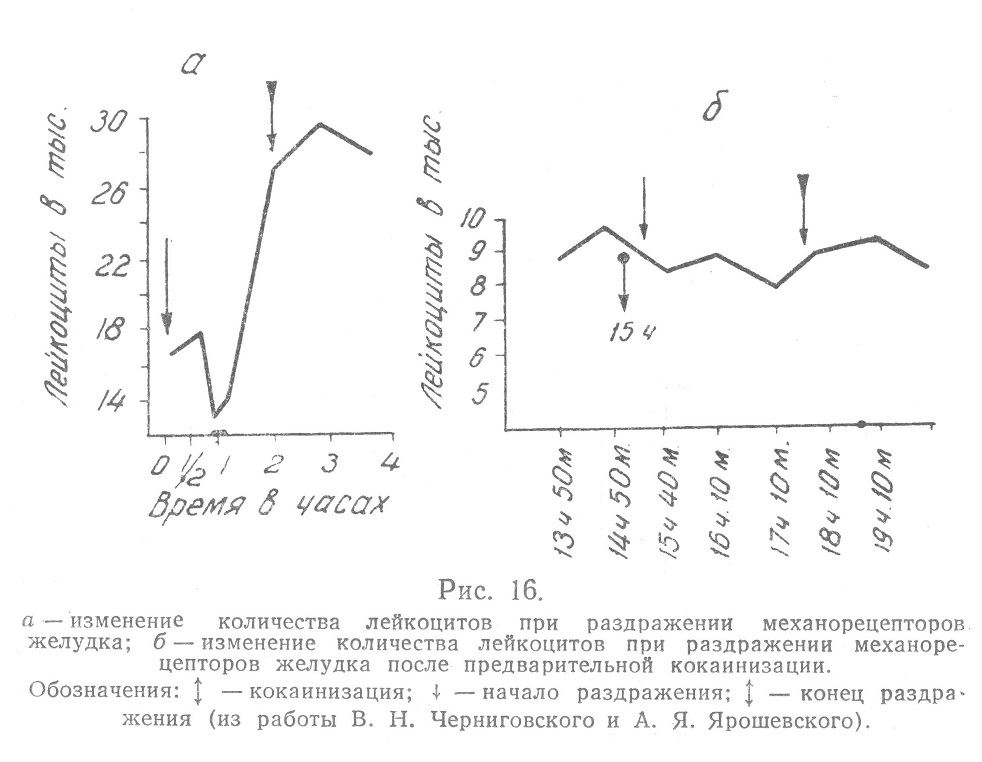
Изменения наблюдались также и в лейкоцитарной формуле, что иллюстрирует приведенная автором табл. 8.
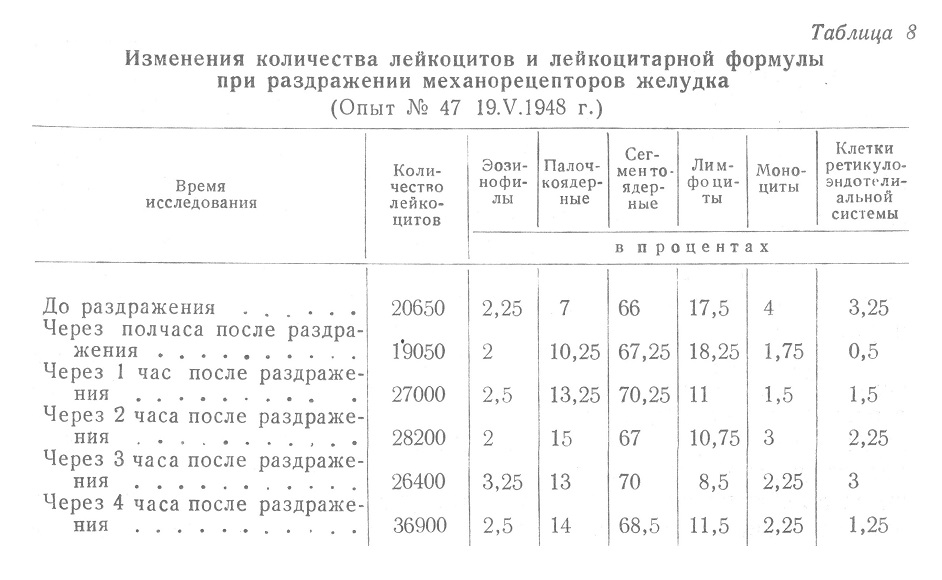
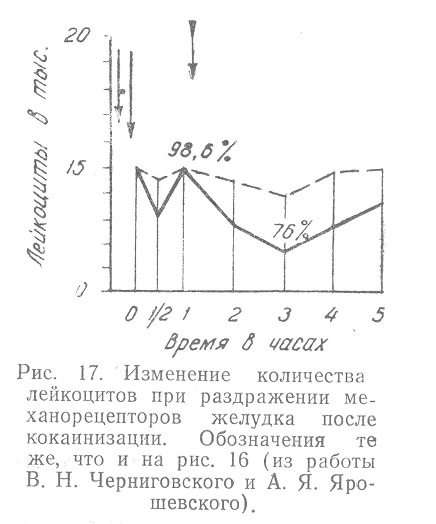
При выключении рецепторов, достигавшемся смазыванием слизистой желудка 2—3% раствором кокаина, изменения количества лейкоцитов после раздражения раздуванием баллона не превышали самопроизвольных колебаний. Указанные изменения представлены на рис. 17.
При постоянном выключении центростремительных путей от желудка, достигавшемся частичной денервацией его, лейкоцитарная реакция в результате раздражения механорецепторов желудка также исчезала. Последующими опытами (В. Н. Черниговский и А. Я. Ярошевский) было доказано, что колебания количества лейкоцитов в крови при раздражении механорецепторов желудка не находятся в прямой связи с выделением желудочного сока. Сдвиги в лейкоцитарной формуле предотвращались при кокаинизации только в 50% случаев, но при раздражении предварительно денервированного желудка закономерные изменения лейкоцитарной формулы не наблюдались уже как правило. Таким образом, был отчетливо установлен рефлекторный характер количественных и качественных изменений лейкоцитарного состава крови, наступающих при механическом раздражении желудка.
Мы привели некоторые данные из многочисленных важных и интересных исследований В. Н. Черниговского и его сотрудников, изложенных довольно подробно в монографии «Вопросы нервной регуляции системы крови» и других работах.
Хотя авторы скромно говорят, что в изучении проблемы интероцептивных влияний на систему крови «сделаны лишь первые шаги» мы должны заметить, что шаги эти достаточно значительны, чтобы дать повод к некоторым далеко идущим, но не всегда обоснованным заключениям, которые воспринимаются часто без достаточной критики и в дальнейшем приводятся уже как твердо установленные положения.
Для примера можно указать на следующее заключение: «Отсутствие полного параллелизма между нарастанием числа эритроцитов и содержанием в периферической крови гемоглобина, а также изменения количества ретикулоцитов (увеличение на 0,5—1%) заставляют нас высказать предположение, что при раздражениях интерорецепторов желудка из костного мозга поступают в периферическую кровь элементы эритроцитарного ряда, менее насыщенные гемоглобином, иными словами, изменения состава крови, которые наблюдаются при интероцептивных раздражениях, происходят при участии костного мозга».
Возникает законный вопрос: о каком же участии костного мозга идет речь — об участии его как органа кроветворения, производящего в ответ на раздражение новые эритробласты, или только как органа депо, участвующего в перераспределении крови? Разъяснение этого вопроса следует из дальнейшего изложения: «...увеличение числа ретикулоцитов, сдвиги в лейкоцитарной формуле, а также довольно часто обнаруживаемые изменения в миэлограмме заставляют нас считать, что интероцептивные раздражения желудка ведут к выбрасыванию из костного мозга форменных элементов крови, а возможно, и к их ускоренному созреванию. Напомним, что образование эритробластов, например, не требует чрезвычайно больших промежутков времени: по данным Дюстина (Р. Dustin) (1950), в одну минуту в организме образуется около двух миллиардов эритробластов».
Иллюстрацией свободного толкования полученных в эксперименте данных может служить табл. 9 упомянутой монографии.
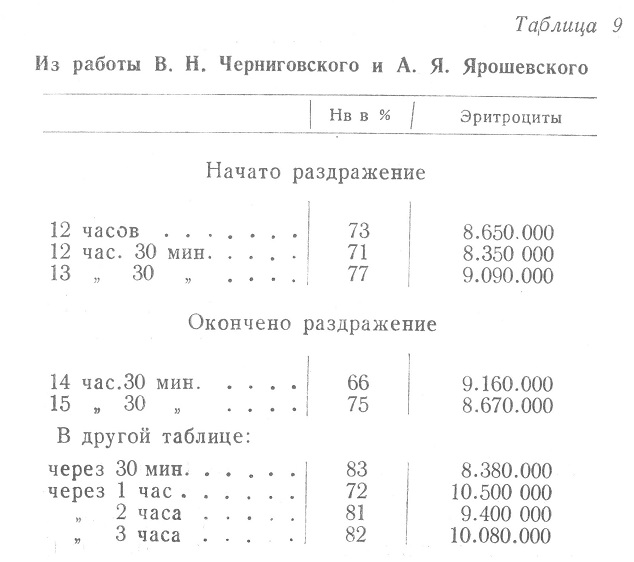
Из данных, приведенных в таблице, следует, что при раздражении механорецепторов желудка наличие гемоглобина возросло максимально На 4%, эритроцитов на 2000000, ретикулоцитов на 0,7%.
Естественно, что сами авторы отмечают такое несоответствие между количеством гемоглобина и эритроцитов и пытаются объяснить его выходом в кровь менее насыщенных гемоглобином эритроцитов костного мозга. Мало того, они считают возможным принять это обстоятельство за доказательство участия костного мозга в изменениях крови при указанном раздражении желудка. Принято, особенно после работ Т. С. Истамановой, считать, что зрелые эритроциты выходят в кровь только как ретикулоциты и в таком состоянии циркулируют в крови 1—2 суток. Это дает возможность учесть диапазон эритропоэза. С другой стороны, известно, что предельное насыщение гемоглобином достигается к моменту пикнотизации ядра эритробласта и с его потерей дальнейшее насыщение гемоглобином не происходит. Следовательно, при физиологических условиях эритроциты костного мозга и крови предельно насыщены гемоглобином. Обращаясь к приведенным в таблице конкретным данным, мы могли бы ожидать, что увеличению количества эритроцитов на 2 000 000 должно было бы соответствовать увеличение гемоглобина примерно на 22%, а не на 3%, как это было в действительности. Если посчитаться с объяснением авторов, то надо было бы признать, что поступившие в ответ на раздражение из костного мозга эритроциты в количестве 2 000 000 гемоглобина почти не содержат. Не говоря уже о невероятности такого вывода, следует напомнить, что количество ретикулоцитов повысилось при этом всего на 0,7%, следовательно, из костного мозга вышло в кровь не 2 000 000, а только (0,7% X 6 700 000) 46 900 эритроцитов в 1 мм3. Если принять, что выпущенные таким образом из костного мозга 46 900 новых эритроцитов совершенно не содержат гемоглобина, то его общее количество должно было бы снизиться всего на 0,5%, а не на 22%. Уже из подсчета ретикулоцитов можно сделать заключение, что 2 000 000 эритроцитов поступили в кровь не из костного мозга, а из других депо.
Дальнейшие исследования тех же авторов с очевидностью показывают, что эритроциты при раздражении желудка поступают, главным образом, из селезенки. Таким образом, очевидно, что далеко идущее заключение авторов об участии костного мозга в изменениях крови при раздражении механорецепторов желудка, выражающееся в «ускорении созревания», нельзя признать правильным.
Дать объяснение самому факту несоответствия между нарастанием количества эритроцитов и гемоглобина, конечно, трудно, но возможно оно кроется в самом методе их определения. Косвенно об этом свидетельствуют данные из других опытов (табл. 9).
Таким образом, резкие колебания с повышением гемоглобина почти на 10% при снижении количества эритроцитов на 1 000 000 и снижение гемоглобина при нарастании эритроцитов в течение часа можно объяснить только неточностью метода исследования.
Несомненный интерес представляют данные исследования пунктатов костного мозга при раздражении механорецепторов желудка. Авторы сами указывают на разнообразные результаты своих опытов, то на нарастание эритробластов, то на их снижение, то на неизменность.
Несмотря на неопределенность результатов, они все же делают вывод, что «видимо изменения в числе эритроцитов при раздражении интерорецепторов желудка осуществляются при участии костного мозга. В тех случаях, когда имело место лишь выбрасывание ретикулоцитов, наблюдалось некоторое уменьшение количества эритробластов третьего порядка. Однако в ряде случаев, наряду с поступлением уже созревших форменных элементов, начинался процесс оживленного созревания эритроцитарного ростка в костном мозгу и количество эритробластов увеличивалось».
Уже неоднократно подчеркивалось, что кроветворение — это сложный процесс, в котором можно отметить три последовательных этапа — размножение, созревание и поступление форменных элементов в кровь. Когда мы говорим о кроветворении, то представляем себе прежде всего первый и основной этап — размножение костномозговых клеток. Вместе с тем ни в одной специальной монографии вопрос о размножении форменных элементов костного мозга при раздражениях не поднимается, и исследования в этом направлении не производятся. Речь обычно идет об ускорении созревания форменных элементов. Но как же можно себе представить ускорение созревания и особенно увеличение количества имеющихся форменных элементов при отсутствии усиления размножения их? Несомненно при этом должен наступить разрыв в последовательности этапов кроветворения и недостаточность последнего. Поскольку так на деле не получается, трудно думать, что в таких случаях имеется стойкое изолированное ускорение созревания.
Вышеприведенное утверждение, что при наличии только выбрасывания эритроцитов уменьшается количество ортохромных эритробластов, не имеет, таким образом, ни теоретических, ни фактических оснований. Рассмотрим некоторые данные из монографии, которые служили, невидимому, поводом В. Н. Черниговскому и А. Я. Ярошевскому для указанных положений (табл. 10).
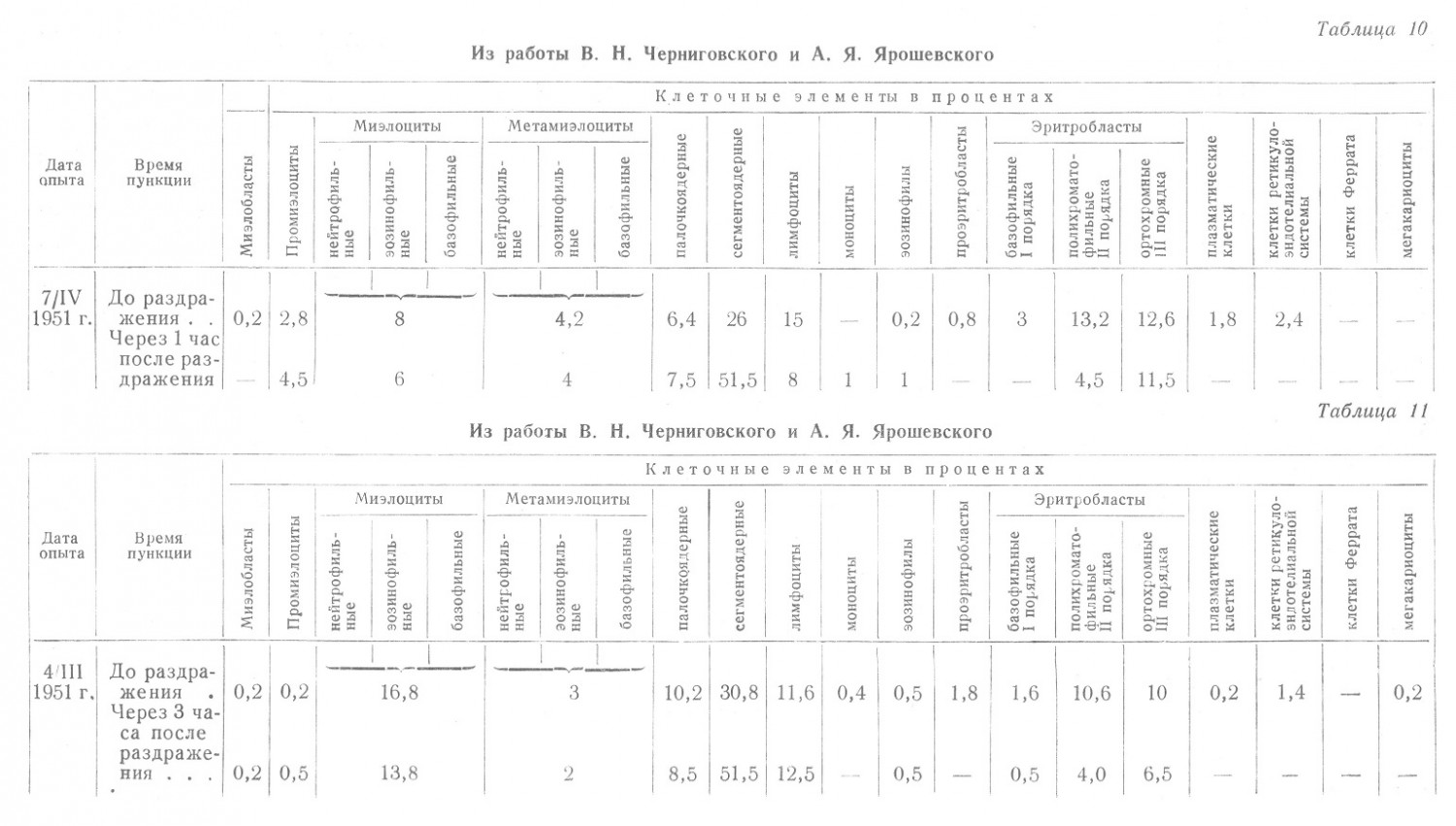
В этом опыте уже через час произошли чрезвычайные, хотя совершенно непонятные сдвиги. Исчезли проэритробласты, базофильные эритробласты, количество полихроматофильных эритробластов уменьшилось втрое, а количество ортохромных осталось почти неизменным. Общее процентное их число с 29,6 снизилось до 16. Однако по кривой созревания можно составить себе представление, что уже через 3 часа после этого картина изменилась.
При отсутствии проэритробластов появляются базофильные эритробласты и втрое возрастает общее число эритробластов. Указанные изменения авторы иллюстрируют в своей работе (рис. 18).
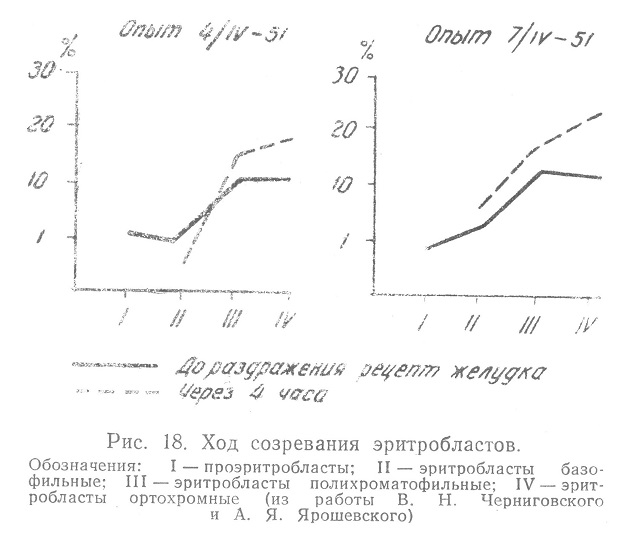
Мы не можем дать сколько-нибудь удовлетворительного объяснения этим фактам, если исключить возможность ошибки в связи с несовершенством метода. В самом деле продолжительность процесса митотического деления составляет в среднем до пяти часов. Если предположить крайнюю степень его ускорения для эритробластов до одного часа, то, чтобы количество эритробластов увеличилось за три часа втрое при одновременном усилении поступления созревающих эритроцитов в кровь, по крайней мере, большую часть эритробластов мы должны были бы видеть в состоянии митотического деления. Такое положение, однако, не наблюдалось.
В другом опыте процентное содержание ортохромных нормобластов действительно снизилось через 4 часа, но при этом снизилось и общее содержание эритробластов с 24% до 11%, т. е. изменения были противоположны вышеприведенным (табл. 11).
И, наконец авторами приведены такие данные (табл. 12 и 13).
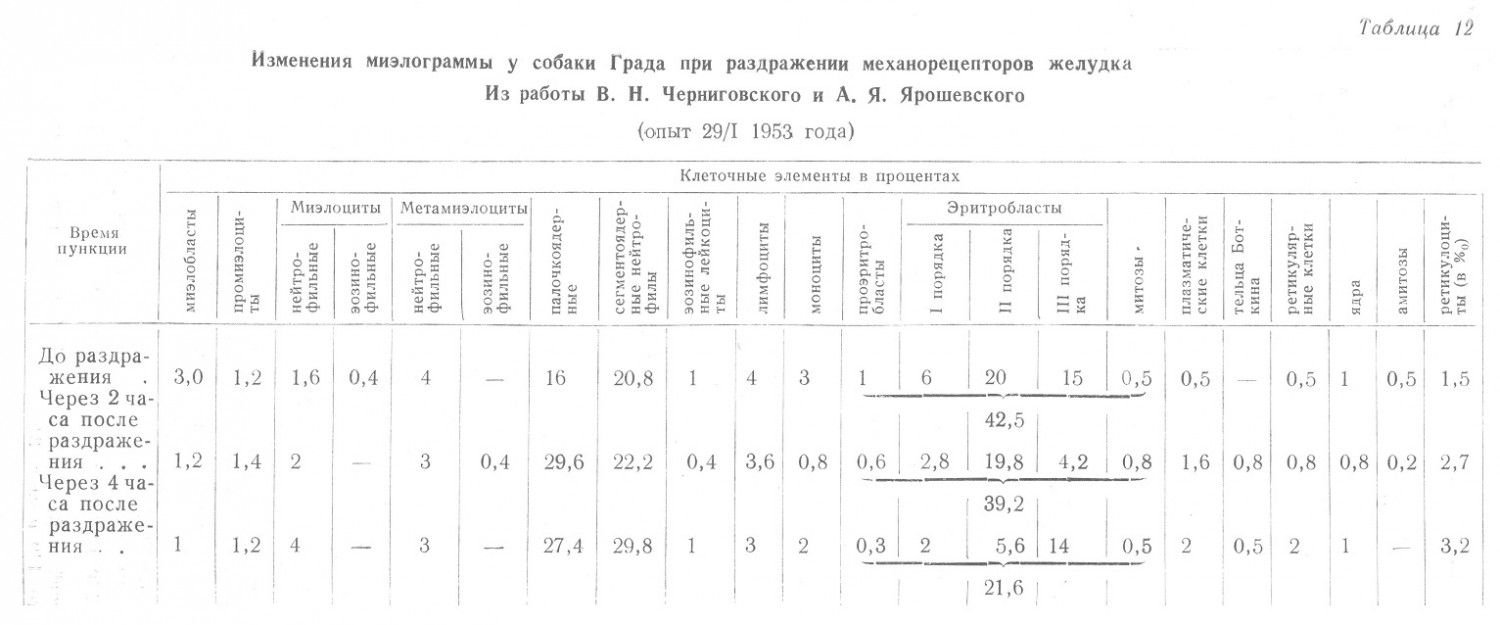
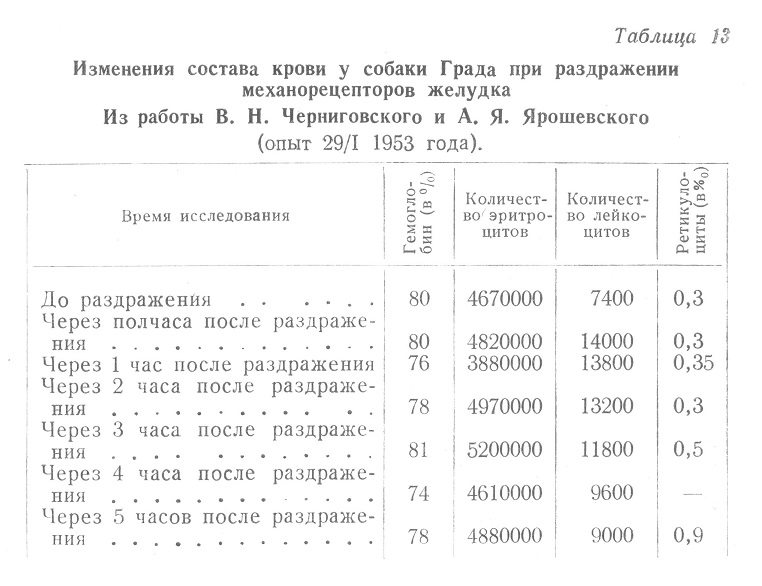
Совершенно очевидно несоответствие между изменениями состава крови и пунктатов костного мозга, причем при значительных изменениях з последнем изменения в крови были незначительными.
Конечно, вопросы соотношения между составом крови и костного мозга как кроветворного органа при раздражении рецепторов других органов требуют дальнейшего изучения, но полученные до настоящего времени данные не дают оснований для выводов об ускоренном созревании форменных элементов при такого рода раздражениях.
Значительный интерес представляют исследования влияния на состав крови денервации отдельных органов. Ряд таких исследований произведен сотрудниками В. Н. Черниговского, исходившими из положения о том, что если раздражение рецепторов влияет на состав крови, то и выключение мощных, а тем более постоянно действующих репецторных зон не может оставаться безразличным для состава крови.
Для изучения изменений, зависящих от выключения афферентной иннервации, были предприняты опыты с денервацией каротидных синусов, обладающих, главным образом, чувствительной иннервацией, и перерезкой депрессорных нервов. После основной операции — денервации каротидных синусов и перерезки депрессорных нервов — вскоре начиналось падение количества гемоглобина, достигавшее максимума между девятым и 14-м днем (в двух из 10 случаев количество гемоглобина снизилось более чем в 2 раза — до 30%), и соответственно снижение эритроцитов в среднем на 2 000 000 в 1 мм3. Цветной показатель существенно не изменялся. Наблюдался выраженный ретикулоцитоз, иногда уже со второго-четвертого дня после оперативного вмешательства, причем по сравнению с исходным уровнем количество ретикулоцитов увеличивалось в 3,5; 4; 4,5; 6; 7 раз. Изменения лейкоцитарного состава крови после этой операции не имели закономерного характера. В. Н. Черниговский и А. Я. Ирошевекий естественно ставят вопрос о том, каков же механизм развития наблюдавшейся анемии. Имели ли они дело с увеличением гемолиза, ответной реакцией костного мозга или с первичным нарушением эритропоэза в костном мозгу?
Авторы считают, что полученные данные позволяют им высказаться в этом отношении лишь предположительно. Правильность постановки вопроса и предупредительно осторожное отношение к выводам несомненно импонирует важности полученных фактов. Тем более удивительными представляются дальнейшие совершенно необоснованные и противоречивые выводы. Уже первое заявление авторов, что «появление большого количества ретикулоцитов в первые же сутки после операции скорее свидетельствует против первичного гемолитического характера анемии», не имеет никаких оснований. Ранее ими указанно, что ретикулоцитоз появляется со второго-четвертого дня и держится на протяжении месяца. Вместе с тем ими же показано, что значительная анемизация также начинается на третий-четвертый день. Однако теоретически и практически хорошо известно и понятно, что прежде чем выявляется при умеренном постепенно нарастающем гемолизе анемия, уже заметен выраженный подъем ретикулоцитов, компенсирующих в первое время усиление кроворазрушения. В клинике нередко наблюдаются случаи, где в начальных стадиях гемолитического синдрома, количество эритроцитов не только не снижается, но при усилении эритропоэтической деятельности костного мозга даже несколько нарастает.
Далее авторы подчеркивают, что ретикулоцитоз и в некоторых случаях даже нормобластоз позволяют отвергнуть гипопластический характер развивающегося малокровия и заканчивают объяснение совершенно неожиданным и неверным заключением: «Следовательно, наиболее вероятно считать, что анемия есть результат нарушения созревания эритробластической ткани и выхода эритроцитов в периферическую кровь».
Клиническими и экспериментальными исследованиями твердо установлено, что при «замедлении созревания эритробластической ткани и выхода эритроцитов в периферическую кровь количество ретикулоцитов снижается. О чем же рассуждают авторы, если указывают, что количество ретикулоцитов возрастало в 4, 6, 7 раз по сравнению с исходными числами? Они не располагали, правда, данными о характере костномозгового кроветворения, но ссылаются на проф. А. В. Соловьева, по данным которого в подобных случаях имелся активный костный мозг, что также противоречит их заключению. Следовательно, все данные в этих случаях позволяют считать наиболее вероятным, что анемия развивается именно в результате преходящего усиления гемолитических процессов.
Это наиболее вероятное заключение соответствует тем данным, которые были приведены в начале главы. Эти данные свидетельствуют о том, что при значительной травме нервной системы (опыты Пенн и других исследователей) так или иначе усиливаются процессы кроворазрушения, а изменения в костном мозгу с активацией эритропоэза являются последующими.
Не станем подробно излагать данные об изменении состава крови, полученью О. И. Моисеевой после денервации печени и Е. И. Кан после денервации селезенки, поскольку они совершенно недостаточны, чтобы на их основе сделать более или менее обоснованное заключение. В. Н. Черниговский и А. Я. Ярошевский, публикующие эти исследования, также воздерживаются от определенных выводов. Вместе с тем они считают «вполне законным предположение, что денервация печени несколько нарушает созревание и, возможно, выход эритробластических (надо полагать эритроцитарных) элементов из костного мозга в периферическую кровь». Косвенным доказательством этого, по их мнению, служат специальные исследования о влиянии наркоза и интероцептивных раздражений на состав крови животных с денервированной печенью, где наблюдалось быстрое поступление в кровь форменных элементов и нарастание содержания гемоглобина и эритроцитов. Вопреки мнению авторов, мы не считаем вышеприведенное положение «вполне законным». Об этом в первую очередь свидетельствуют только что изложенные факты полной возможности быстрого поступления форменных элементов в кровь, не позволяющие предположить замедление созревания, иначе нельзя было бы допустить специально подчеркнутую авторами «быстроту» поступления. Кроме того, об этом свидетельствует полное отсутствие изменений в составе пунктатов костного мозга до и после операции (табл. 14).
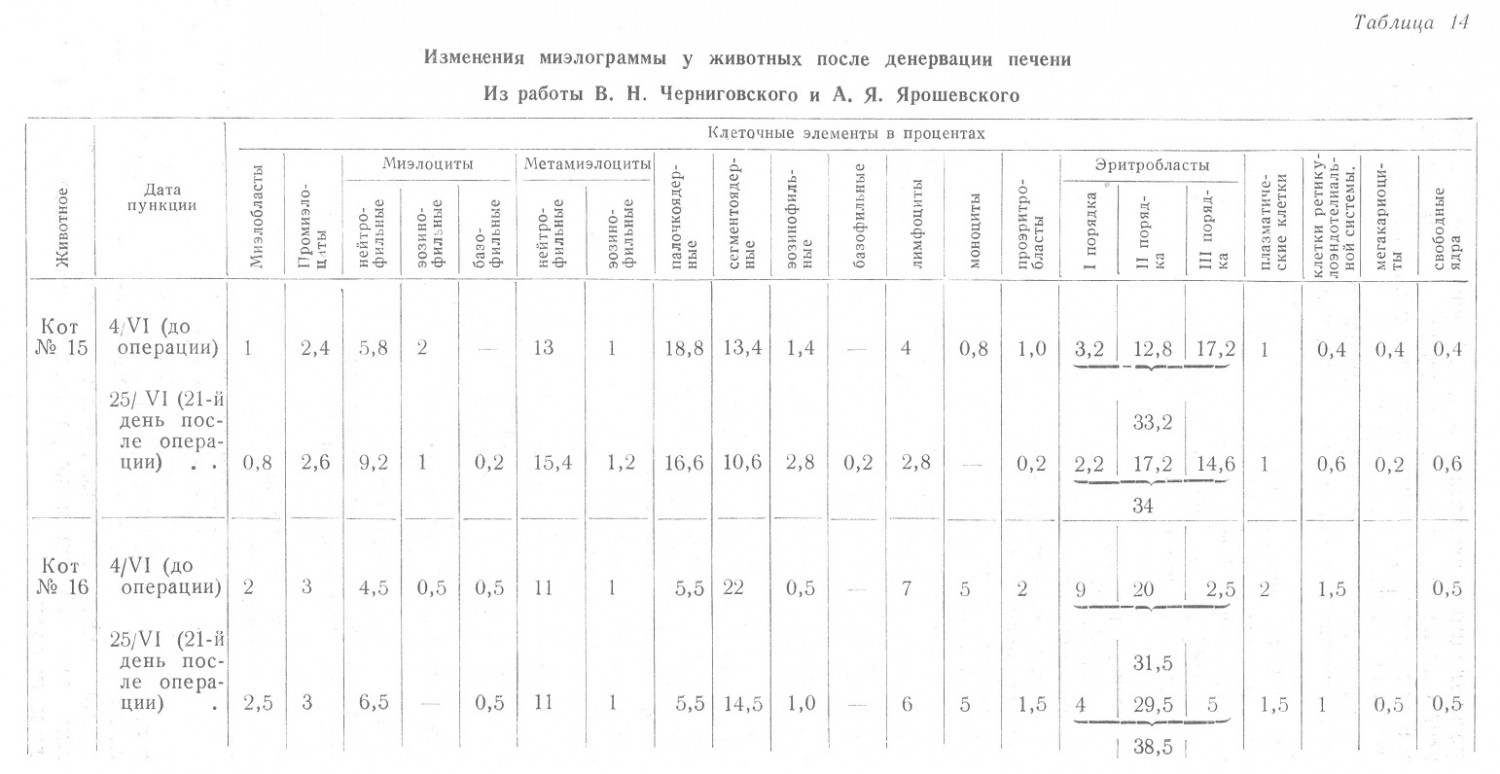
При отсутствии изменений ретикулоцитов нельзя предполагать и сколько-нибудь выраженный гемолиз. Следовательно, на основании имеющихся данных, единственно законным является предположение, что быстрое развитие анемии и есть результат депонирования значительного количества крови и, в частности, эритроцитов в печени, а возможно и в других органах брюшной полости. Нет сомнения в том, что дальнейшие исследования прольют свет на рассматриваемые, еще не вполне выясненные, вопросы, но до этого времени следует воздержаться от утверждения того, что «денервация печени влияет на кроветворение» и что «кроветворная функция» печени находится под контролем нервной системы, так как известно, что в условиях физиологии кроветворение в печени у взрослого животного не происходит и, следовательно, кроветворной функцией она не обладает.
В качестве другой модели для изучения значения афферентной иннервации в регуляции состава крови тем же авторам служила лейкоцитарная реакция, обусловливаемая внутримышечным введением молока. При этом в течение первых 0,5—1 часа наблюдалось снижение количества лейкоцитов, которое через 1—2 часа сменялось лейкоцитозом. При предварительной новокаинизации места введения молока в течение четырех-семи часов лейкоцитоз не развивался, а лейкопеническая фаза длилась до пяти часов.
Контрольное введение 1 % раствора новокаина вело только к выраженной лейкопении в течение трех-четырех часов. При одновременном введении новокаина и молока в различные места лейкоцитарная реакция соответствовала обычной для введения молока.
При введении трех миллилитров молока в мышцы бедра деафферентированной конечности наблюдались изменения в содержании лейкоцитов, близкие к изменениям при введении молока после предварительной новокаинизации.
Заслуживает внимания факт, что во всех этих опытах на качественные изменения состава лейкоцитов ни предварительная новокаинизация, ни деафферентация влияния не оказывали и ядерный сдвиг в лейкоцитарной формуле не только сохранялся, но становился даже несколько более выраженным.
Результаты, подобные вышеприведенным, получены М. Д. Школьниковой при введении в мышцу кошкам ослабленной культуры туберкулезной палочки (БЦЖ). У животных, находившихся под нембуталовым наркозом лейкоцитоз не наступал, но качественные изменения с ядерным сдвигом влево сохранялись. На основании приведенных данных В. Н. Черниговский и А. Я- Ярошевский заключают, что «вывод, который можно сделать на основании этих опытов, совершенно ясен. Очевидно, лейкоцитарная реакция находится в прямой связи с тем, что раздражитель на первых же порах встречается с нервной системой, именно ее периферической воспринимающей частью и рефлекторно вызывает те или иные изменения состава крови. Вместе с тем мы не можем отрицать и значения гуморальных факторов, так как качественные изменения состава крови в виде резкого сдвига лейкоцитарной формулы влево сохраняются и в тех случаях, когда произведена полная деафферентация конечности».
С таким выводом нельзя как будто не согласиться. Вместе с тем не все и здесь представляется вполне ясным. В самом деле новокаин должен снять рефлекторную лейкоцитарную реакцию, в которой несомненно наблюдаются две фазы — лейкоцитоз и предшествующее ему снижение количества лейкоцитов. На деле же снижение количества лейкоцитов сохраняется или становится даже более выраженным, во всяком случае в смысле продолжительности. Такое снижение можно было бы отнести за счет влияния самого новокаина, как это следует из контрольных опытов, но затруднительным тогда остается объяснение фазы снижения количества лейкоцитов при введении молока в деафферентированную конечность. По-видимому, сама рефлекторная реакция сложнее, чем это представляется, и к тому же она несомненно имеет перераспределительный характер, о чем и свидетельствует первая фаза, в полу- или часовое течение которой количество лейкоцитов может снизиться вдвое. Так же очевидно, что гуморальное влияние молока происходит независимо от рефлекторной реакции и, следовательно, в приведенных опытах, как и в большинстве других, изменения состава крови определяются сочетанием перераспределительной реакции, подчиненной непосредственно нервной системе и реакции со стороны кроветворного органа, которая по крайней мере в приведенных примерах протекает независимо от влияния нервной системы.
По поводу основной задачи опытов с денервацией каротидных синусов — показать роль афферентной и эфферентной иннервации органов в регуляции состава крови, сами авторы заключают: «Происходят ли сдвиги в составе крови в результате только выпадения рефлекторных влияний с синокаротидной зоны, мы сказать не можем. После денервации кародитного синуса в организме наступает ряд изменений обмена, в том числе и минерального, изменения гемодинамики и др.; все это, конечно, может оказать влияние на состав крови. В этом направлении требуются дальнейшие исследования».
Однако, если при этом изменения обмена, гемодинамики и др. оказывают влияние на состав крови, то, естественно, значение выпадения рефлекторых влияний и тем самым роль афферентной иннервации в регуляции состава крови значительно умаляется. Приведенные опыты, позволившие установить факт изменения состава крови, не проливают свет на механизм этих изменений и роль эфферентной иннервации. Несомненно также, что из этих опытов не вытекает регулирующее влияние нервной системы на кроветворный орган (костный мозг) и кроветворение. В этом отношении заслуживают внимания исследования М. Л. Беленького и Ю. Н. Стройкова, изучавших роль каротидных клубочков в развитии эритроцитоза при тканевой гипоксии, вызываемой воздействием цианидов.
Чтобы выяснить роль каротидных химиорецепторов в такой реакции, авторы произвели «фармакологическую денервацию» каротидных клубочков путем инфильтрирования области каротидных синусов с обеих сторон 1% раствором новокаина. Новокаинизация синусов резко ослабляла, снимала или даже извращала реакцию со стороны крови на внутривенное введение цианида.
Для того, чтобы убедиться в существовании рефлекторной регуляции содержания эритроцитов, авторы исследовали состав крови при перфузии изолированного каротидного синуса растворами цианида. При этом во всех случаях увеличивалось количество эритроцитов в периферической крови. Доказав, таким образом, роль рефлексов химиорецепторов каротидного синуса в изменениях крови при острой тканевой гипоксии, авторы, предполагая перераспределительный характер изменений состава крови, повторили эти опыты на животных после удаления у них селезенки. Спленэктомия во всех случаях исключает рефлекторный эритроцитов, причем наблюдается даже тенденция к обратному эффекту, т. е. к снижению количества эритроцитов. Единственной причиной рефлекторного эритроцитоза при острой гипоксии эти исследователи считают выбрасывание из селезенки в кровь депонированных в ней эритроцитов. Предположение о том, что сокращение селезенки является результатом рефлекторного возбуждения симпатических волокон, иннервирующих этот орган, они подтверждают тем, что внутривенное введение симпатолитина исключает или даже извращает изменение количественного состава эритроцитов. В этих опытах М. Л. Беленького и Ю. Н. Стройкова очевиден и перераспределительный характер изменений состава крови и механизм их развития. Не исключается возможность, что денервация каротидного клубочка изменяет состав крови тем же путем, воздействуя на селезенку как орган кроворазрушения, т. е. оказывая влияние на гемолитическую ее функцию.
В конце концов и В. Н. Черниговский и А. Я. Ярошевский, которым принадлежит наиболее обширная часть исследований роли нервной системы в регуляции состава крови, приходят к заключению: «Мы не можем, конечно, утверждать, что при денервации ряда органов изменения в органах системы крови зависят лишь от выключения афферентной импульсации. Наоборот, мы убеждены, что при денервации печени и селезенки имеет место и значительное нарушение их функций с последующим влиянием на состав крови».
Другой вопрос, который уместно здесь обсудить, касается существования специальных центров кроветворения и кроверазрушения. Гипотеза о существовании центров была высказана С. П. Боткиным. Последующие работы Розенова (Rosenow), Гофа и Ленгардта, Шульгофа (Schulhoft) и Матиеса (Mathies), Гейльмейера и других авторов дали повод многим исследователям признать существование в межуточном мозгу центров кроворазрушения и кроветворения, даже раздельно центров эритро- и лейкопоэза. В 1950 г. мы высказали сомнение в существовании таких обособленных центров, что вызвало возражения, а у некоторых неадэкватную реакцию. Так, М. С. Дульцин по этому поводу писал: «несколько легкомысленно толкуется автором вопрос о центрах кроветворения. Читателю преподносится следующее заключение: «...центры кроветворения очень уж проблематичны. В связи с этим уместно напомнить замечание В. Н. Черниговского, что указанная проблема разрешена И. П. Павловым — «центрами» кроветворения является кора головного мозга». Для сведения читателей и М. С. Дульцина в том числе, надо подчеркнуть, что И. П. Павлов никогда проблемой кроветворения и его центров не занимался и потому этой проблемы не мог и разрешить.
Полагаем, что В. Н. Черниговский не нуждается в защите справедливости тех или иных его положений, но все же мы должны усомниться в том, что такое сугубо неверное положение было действительно им высказано. Нам кажется, что даже мало искушенному читателю ясно, что кора головного мозга имеет более широкое назначение, чем служить центром кроветворения.
Не об этом, по-видимому, говорил и В. Н. Черниговский. В статье «Современные проблемы физиологии интерорецепторов» он писал: «Между тем для признания влияния нервной системы на систему крови нет надобности говорить об особом высшем центре, расположенном з центральной нервной системе. Искать этот центр незачем. Он давно найден И. П. Павловым и называется корой больших полушарий головного мозга, которой подчинена регуляция всех процессов в организме». В монографии 1953 г. В. Н. Черниговский и А. Я. Ярошевский пишут: «Не существует особых специфических рефлексов, регулирующих состав крови и имеющих в центральной нервной системе свои четко обособленные центры, свою строгую локализацию». Некоторые авторитетные ученые сомневаются в существовании не только центра кроветворения, но и других гипотетических центров. Так, Б. И. Баяндуров пишет: «Подобно А. А. Богомольцу и др., мы также отрицаем существование в межуточном мозгу каких-либо специфических центров обмена веществ (углеводного, белкового, водно-солевого и др.) равно как и специального центра теплорегуляции. Мы разделяем известный взгляд о том, что в межуточном мозгу находятся высшие симпатические и парасимпатический центры».
Против возможности признать в настоящее время точку зрения морфологической локализованности центров кроветворения высказывается и В. Г. Вогралик. По поводу изменений состава крови при энцефалографии, которые принимаются как особенно доказательные для существования центров кроветворения, Т. С. Истаманова высказывает мнение, что они являются следствием раздражения сосудодвигательного центра, а не специального центра кроветворения.
Сомнения в существовании особых центров кроветворения оправданы тем более, что до настоящего времени не представлены доказательства непосредственной нервной регуляции кроветворения, т. е. размножения и созревания кровяных элементов в кроветворных органах. Выяснение этого последнего положения и является собственно основным и наиболее актуальным.
В связи с развитием физиологического учения И. П. Павлова в настоящее время едва ли нужно сомневаться в том, что все воздействия внешней или внутренней среды организма высшего животного или человека, вызывающие со стороны этого организма те или иные ответные реакции, происходят при непременном и обязательном участии нервной системы (А. Г. Иванов-Смоленский).
Значит ли это, что всякое участие нервной системы должно выражаться в регулирующем влиянии, что при признании для целостного организма человека даже полного преобладания центральной нервной регуляции над ее подчиненной гуморальной мы должны отказаться совершенно от представления о возможности сохранения гуморальной, химической регуляции для отдельных процессов, в частности, в органах кроветворения в связи с особенностью их функции, и всякое гуморальное влияние должны рассматривать только как звено в центральной нервной регуляции, если до настоящего времени это для процессов кроветворения не удалось подтвердить клинически и экспериментально? Значит ли это, далее, что мы должны усомниться в существовании или значении автоматических раздражителей и отказаться от права полагать, что в организме могут существовать, кроме коры головного мозга и в других органах, клеточные элементы с особой чувствительностью к прямым воздействиям определенных химических раздражителей, влияющих на их функцию? При этом нет нужды отрицать возможность влияния нервной системы на другие стороны их жизнедеятельности, например, на их трофику.
Нельзя не считаться с тем, что в опытах с парэнтеральным введением молока при предварительном воздействии новокаином или при денервации качественные изменения состава крови, зависящие от функциональной деятельности кроветворных органов, сохраняются и все исследователи признают здесь гуморальное влияние. Другое дело, что не все учитывают, что в общей реакции на введение молока надо различать не две части одной и той же реакции, а две реакции со стороны двух систем — сосудистой и кроветворения, т. е. одну перераспределительную, а другую кроветворную.
Известно, что в приспособлении организма к кислородному голоданию значительную роль играет увеличение количества эритроцитов как за счет перераспределительных реакций, так и за счет усиления эритропоэтической деятельности костного мозга. Известно также, что недостаток кислорода является автоматическим раздражителем центральной нервной системы, которая мобилизует соответствующие компенсаторные механизмы.
Большинство исследователей представляет себе, что недостаток кислорода является также непосредственным стимулом к раздражению эритробластов и усилению эритропоэза.
Существуют и другие мнения, в частности, мнение о рефлекторном характере влияния гипоксии. Так, В. Н. Черниговский пишет: «Другие — недостаток кислорода, избыток углекислоты, pH, цианистые соединения раздражают рецепторы не непосредственно, а путем изменения обмена веществ в самих клетках ткани. Изменения же обмена служат прямым раздражителем рецепторов». Следовательно, при недостатке кислорода произойдет, особенно в чувствительном к этому эритробласте, изменение обмена, которое не должно влиять на биологию и функцию этой клетки, но должно повлиять на рецепторы, и только в результате сложного рефлекторного процесса, должного в свою очередь оказать влияние на обмен клетки, наступит соответствующее изменение в биологии и функции эритробласта.
Нам такое мнение не представляется убедительным и, если недостаток кислорода приводит к изменению обмена в клетке, то нам кажется излишним и искусственным привлечение к этому еще рецепторов. Более убедительно выглядит представление о том, что легкая степень гипоксии является достаточным раздражителем только особо чувствительных к кислородному голоданию элементов центральной нервной системы, которая приводит в действие компенсаторные механизмы со стороны сердечно-сосудистой и дыхательной систем. Более выраженная степень гипоксии обусловливает уже некоторое раздражение также высоко чувствительных к недостатку кислорода эритробластов и, таким образом, помимо нервной системы, в компенсацию включается кроветворная система с развитием эритропоэза. Только при еще более выраженной гипоксии и недостаточности компенсаторных механизмов изменяется жизнедеятельность других тканей и органов.
О регуляции кроветворения, часть третья
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
|
