Первое полное описание непрямого деления растительных клеток, как уже упоминалось, мы находим у нашего соотечественника И. Д. Чистякова в его работе «Материалы к истории растительной клетки», вышедшей в 1874 г.
Данные, полученные И. Д. Чистяковым на живем объекте (развитие спор у плауна и хвоща), иллюстрируются прекрасными рисунками. Его исследования представляют большой интерес как первые точные описания кариокинетического деления, притом сделанные на живых объектах.
Одним из первых исследователей кариокинетического деления животных клеток (на фиксированном материале) был русский гистолог П. И. Перемежко (1879).
Сложные морфологические изменения клетки, получившие название кариокинеза, наступают после более или менее длительного периода развития клетки—интеркинеза, который получил неправильное название периода «покоя». В действительности ни о каком покое живой клетки не может быть и речи: она непрерывно изменяется в процессе обмена веществ. Интеркинез — это тот период, когда клетка живет, самообновляется. В течение этого периода развития клетки мелкие незаметные изменения, накопляясь, приводят к ряду видимых изменений: меняется вязкость цитоплазмы, характер ее движения, электрический потенциал, показатель преломления и пр.; изменяется и вся внутриклеточная морфология. Таким образом, в результате развития клетки вслед за интеркинезом наступает период резких видимых изменений, которые свидетельствуют о глубоких внутренних превращениях клетки. Эти превращения обмена всей клетки выражаются в ряде следующих друг за другом морфологических изменений, заканчивающихся возникновением вместо одной клетки двух клеток, которые снова переходят к стадии интеркинеза, или «покоя».
Обмен веществ вновь возникших молодых клеток и сходен с обменом веществ материнской клетки, и отличен от него. Он сходен постольку, поскольку вновь возникшие клетки осуществляют процесс самообновления, т. е. строят из элементов окружающей среды свое собственное тело, развившееся из тела материнской клетки и потому с ней сходное. Они не сходны с материнской клеткой, поскольку они — новые клетки, поскольку в построении тела из элементов окружающей среды всегда возникает не тождественное, а более или менее отличное от материнского тела.
Название «кариокинез» так же неправильно, как и название стадии «покоя», так как оно отражает изменение не всей клетки, а только ядра. Объясняется это неправильное название тем, что изучение клетки, в частности, кариокинетического деления, велось главным образом на фиксированных и окрашенных препаратах, а такая методика, как указывалось выше, позволяет особенно хорошо исследовать именно ядро клетки. Глубокие же изменения, происходящие в цитоплазме, обычно ускользают от наблюдения.
По-видимому, этим обстоятельством (значительно более интенсивной окрашиваемостью ядра) отчасти объясняется и то исключительное внимание, которое в течение ряда десятилетий уделялось изучению ядра и привело морганистов к концепции монопольной роли ядра в передаче наследственных свойств от родителей к детям. Эти глубоко ложные представления, как будет показано ниже, не соответствуют фактическим данным.
Весь процесс непрямого деления клетки условно разделяют на четыре периода, или стадии: профазу, метафазу, анафазу и телофазу (рис.). Каждая предшествующая стадия подготовляет, обусловливает последующую.
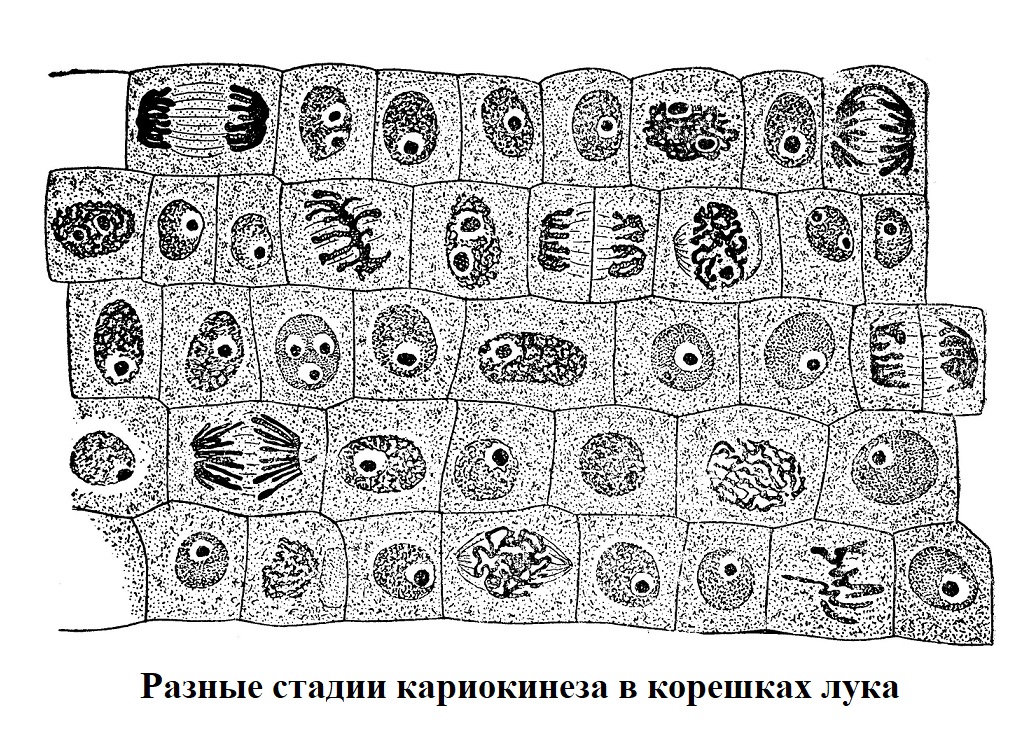
Во время профазы ядро набухает, хроматин (окрашивающееся вещество ядра) приобретает форму сплошной изогнутой нити — плотного клубка; ядрышко исчезает, клубок постепенно разрыхляется и хроматиновая нить распадается на отрезки — хромосомы. В конце профазы ядро как таковое перестает существовать.
На следующей стадии, в метафазе — хромосомы, до того разбросанные беспорядочно, располагаются в средней части клетки более или менее в одной плоскости, образуя экваториальную пластинку, или материнскую звезду. Последнее наименование связано с тем, что хромосомы, если рассматривать их со стороны одного из клеточных полюсов, образуют фигуру, напоминающую звезду. На стадии метафазы обычно и производится изучение морфологии хромосом и их подсчеты. Отдельные хромосомы могут иметь в одной и той же клетке разную форму и величину; они могут отличаться и по своему тонкому строению.
Различны и наборы хромосом у разных животных и растений. Хромосомы то имеют вид прямых и изогнутых палочек, то более или менее округлых зерен. В метафазе происходит продольное деление хромосом, так что общее количество их удваивается.
Во время анафазы каждая из дочерних хромосом отходит к противоположному полюсу клетки, где они и собираются, образуя дочерние звезды.
В телофазе из дочерних хромосом формируется два ядра. Одновременно с этим тело клетки разделяется надвое.
В животных клетках они делятся путем перешнуровывания, в растительных же клетках образуется новая перегородка, разгораживающая старую клетку на две части.
Однако ошибочно было бы думать, что изменения во время кариокинеза ограничиваются только одним ядром и передвижениями хромосом. В действительности при этом происходят глубочайшие изменения всей клетки.
Для кариокинеза, помимо образования хромосом, характерно также формирование ахроматиновой фигуры, в тесной связи с которой происходят все перемещения хромосом. Она представляет собой веретенообразную, волокнистую структуру, расположенную вдоль по длине клетки, в её центральной части.
В клетках, обладающих центросомами, последние лежат на противоположных, заостренных концах веретена и могут быть окружены лучистым сиянием. Вопрос о происхождении веретена не совсем ясен. Вероятно, оно может быть различной природы, — цитоплазменной и ядерной.
Во время клеточного деления происходят существенные изменения всех клеточных органоидов. Центросомы расходятся к противоположным полюсам клетки. Внутриклеточный аппарат, по данным Д. И. Дейнеки, распадается на отдельности — диктиосомы, которые сначала рассеяны по всей цитоплазме, а затем, в анафазе, обычно собираются в две группы вокруг центросом. По окончании деления из диктиосом вновь формируются сети. Хондриосомы в ряде случаев располагаются в экваториальной области клетки и при ее перешнуровывании делятся между дочерними клетками. На других объектах наблюдалось скопление хондриосом на полюсах клетки во время анафазы.
В делящейся клетке резко изменяются все физические и физико-химические показатели: показатель преломления, вязкость, поверхностное натяжение, кислотность и т. д. Эти изменения носят различный характер в разных фазах кариокинеза (так, снижение вязкости может сменяться ее повышением и т. д.).
Новейшие наблюдения на живом материале открывают новые, важные для понимания клеточного деления стороны. Особенно хорошо выявляются изменения делящейся клетки при исследовании ее путем микрокиносъемки. Благодаря особым приемам съемки, можно на экране проследить во всех деталях деление живой клетки. Советским цитологом В. И. Фаворским был заснят фильм деления клеток волосков тычиночных нитей традесканции. Волоски эти состоят из одного ряда прозрачных клеток, что позволяет легко наблюдать под микроскопом происходящие в них изменения, не повреждая клеток. Эти клетки представляют собой одни из классических объектов для демонстрации движения цитоплазмы. Спокойное, струящееся движение, свойственное клеткам этих волосков в стадии «покоя», прекращается во время деления. Движение цитоплазмы во время кариокинеза напоминает движение жидкости во время кипения.
Отсюда мы заключаем о глубоких превращениях, об изменениях обмена веществ клетки. Когда дочерние клетки оказываются сформированными, возобновляется прежнее спокойное, струящееся движение цитоплазмы.
Во время деления и на живом видна фигура веретена, однако нитей, хорошо заметных на фиксированном материале, на живом наблюдать не удалось: возможно, что эти нити являются результатом искусственного воздействия реактивов, т. е. артефактом.
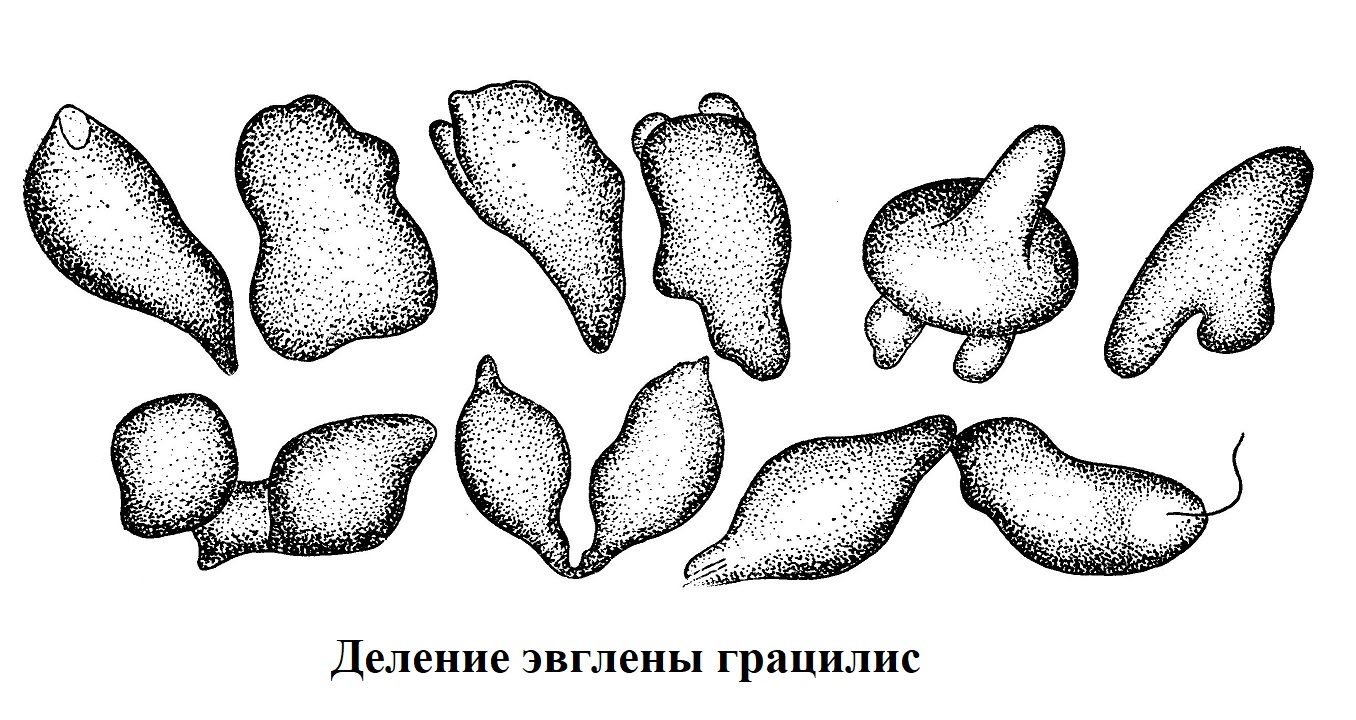
Во время кариокинеза животные клетки округляются, начинают сильно преломлять свет. В анафазе и особенно в телофазе, как показали микрофильмы, снятые с живых объектов, происходит как бы вскипание цитоплазмы, появление и быстрое исчезновение в ней вакуолей. На поверхности клетки образуются многочисленные выросты. Подобные же изменения движения и формы во время деления можно видеть у некоторых простейших.
На рис. изображено несколько этапов деления эвглены, сопровождающегося резким изменением характера движения: поступательное движение организмов останавливается и наступает непрекращающееся во время всего процесса деления энергичное перемещение, внутреннее течение цитоплазмы, которое связано со значительными изменениями формы тела.
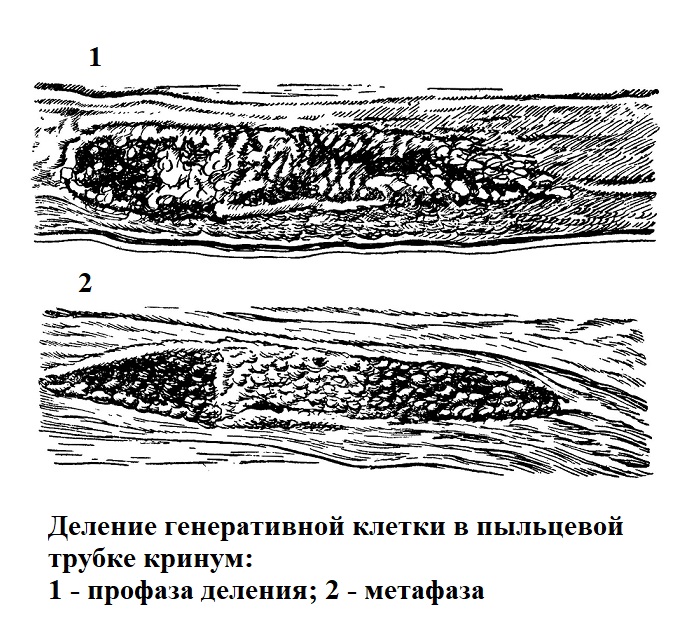
В растительных клетках, не имеющих твердой оболочки, например, генеративной клетке пыльцевой трубки покрытосемянных, также можно наблюдать изменения формы и характера движения в процессе деления (рис. 46).
Все это говорит о том, что кариокинетическое деление представляет собой критическое состояние, охватывающее клетку в целом, а не только ядро.
Несколько подробнее необходимо остановиться на образовании хромосом, поскольку этот вопрос тесно связан с критикой хромосомной теории наследственности.
Исследования, произведенные М. В. Чернояровым и П. В. Макаровым, показали, что хромосомы возникают в каждом кариокинезе заново. Процесс образования хромосом — одно из проявлений присущей каждой клетке способности к формообразованию, как проявлению обмена веществ. Так, в самой ранней профазе в однородном микроскопически ядре появляются отдельные мелкие зернышки, возникающие из субмикроскопической области. Количество их со временем увеличивается. Затем эти зернышки начинают соединяться, сливаться в более крупные глыбки, из объединения которых и формируются хромосомы (рис.).
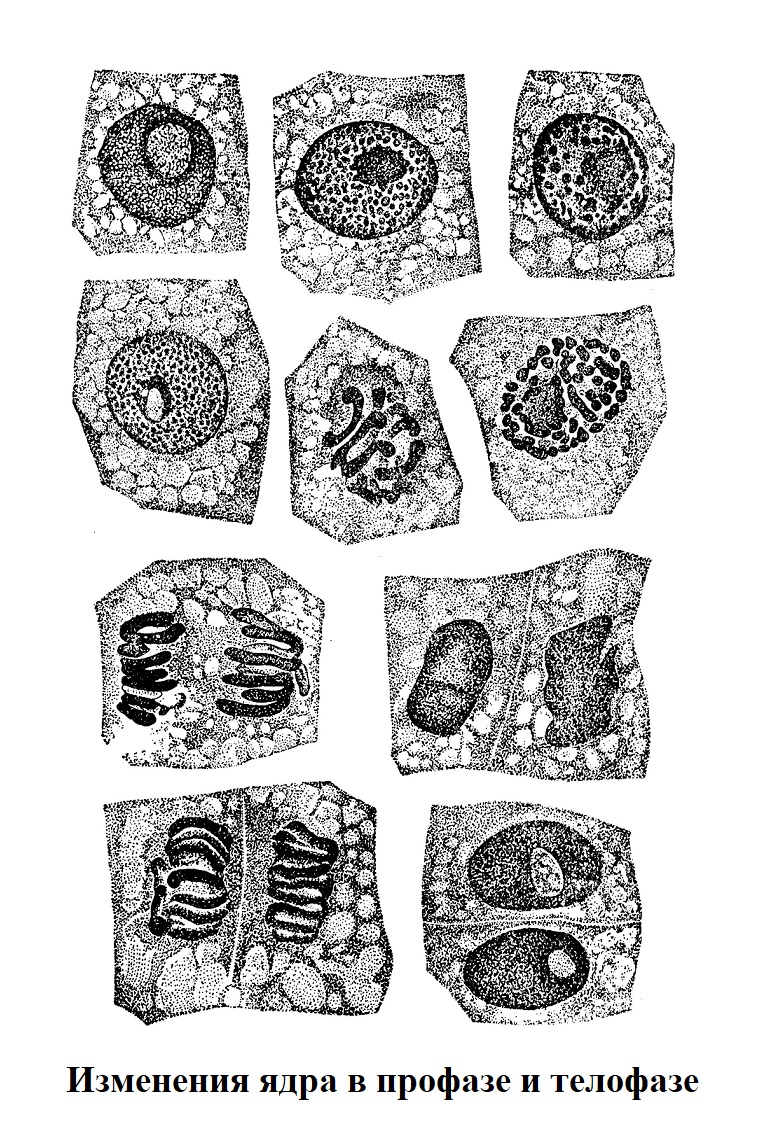
Хромосомы, возникающие в начале кариокинеза, в телофазе снова исчезают, дав начало изменчивому, подверженному обменным превращениям ядерному веществу. Следовательно, хромосомы — не постоянные клеточные структуры, а временные образования, возникающие во время клеточного деления.
Мы видим, что деление клетки сопровождается рядом превращений, свидетельствующих о глубоких и длительных изменениях всего ее обмена веществ. Одно из морфологических проявлений этих перемен и представляют те постепенно развертывающиеся изменения, которые мы, описывая кариокинез, разбили на отдельные этапы—профазу,метафазу, анафазу и телофазу.
Вейсманисты — морганисты защищали так называемые «законы» постоянства числа и формы хромосом. Число хромосом будто бы строго постоянно: оно специфично для организмов данного вида и не подвергается изменениям. В действительности же можно говорить лишь об относительном постоянстве числа хромосом, обусловленном относительной константностью тех условий, при которых происходит их формирование.
Против этого «закона» еще в 1923 г. поднял свой голос И. В. Мичурин. Он писал: «Нельзя быть уверенным в возможности предварительного определения в каждом сеянце-гибриде количества клеточных хромосом, так как таковое (т. е. количество) в гибридных сеянцах плодовых деревьев не только не одинаково во всех частях одной и той же особи, но количество это не постоянно в своей величине и проявлении в течение всего периода времени, требующегося для полной формировки строения растения каждого сеянца-гибрида, что иногда заканчивается в продолжении десятков лет». Мичурин и в этом был прав. Последующее исследование развеяли мифический «закон» морганистов.
Уже давно накапливался материал, доказывающий изменчивость числа хромосом, притом не только в клетках разных организмов одного вида, но даже в разных тканях и органах одного и того же организма и, наконец, в разных клетках одной и той же ткани. Такой материал в значительном количестве был получен при исследованиях как на растительных, так и животных объектах. Даже на таком хорошо изученном объекте, как человек, у которого, казалось бы, прочно установлено постоянное наличие 48 хромосом при делении всех клеток тела (кроме гамет), отмечены большие колебания числа хромосом; особенно значительные отличия обнаружены в тканях эмбрионов, где число хромосом в метафазе деления колеблется от 24 до 64. В тканях взрослых нормальных людей тоже нередки колебания числа хромосом от 40 до 58. Эти и многие другие данные, полученные при цитологическом исследовании как животных, так и растительных объектов, свидетельствуют о том, что клетки организма на разных этапах его развития не тождественны.
Для иллюстрации того, как мало характерна эта отдельно взятая черта — количество хромосом, — можно привести такие примеры: 16 хромосом имеется у коровы, морской свинки, лука; 24 хромосомы — у мыши, саламандры, лимона и т. д.
Форма хромосом тоже лишь относительно постоянна. При изменении внешних условий, например, при понижении температуры, форма хромосом резко изменяется. То же наблюдается и в одних и тех же клетках, находящихся в разных стадиях развития. При этом нити превращаются в зерна, зерна — снова в нити и т. д.
Следовательно, нет оснований говорить о постоянстве числа и формы хромосом, как о чем-то абсолютном и раз навсегда данном. Как и другие признаки, они могут изменяться.
Неопровержимое доказательство изменчивости числа и формы хромосом и одновременно новообразования последних дают опыты по вегетативной гибридизации. Работами И. Е. Глущенко, Т. Ф. Поляковой, Г. Б. Медведевой установлено, что в семенном поколении вегетативных гибридов изменяется как число, так и форма хромосом по сравнению с исходной формой. О том же свидетельствуют опыты Т. Д. Лысенко по превращению под влиянием условий развития 28-хромосомных твердых пшениц в 42-хромосомные мягкие, а также возможность превращения пшеницы в рожь. «В определенных условиях жизни, — пишет Т. Д. Лысенко, — в растениях пшеницы происходят незаметные, скрытые, постепенные количественные изменения, которые приводят к быстрым, внезапным, открытым превращениям отдельных клеток растения, в данном случае яйцеклеток пшеничного растения, в качества другого вида. Вместо клетки пшеницы возникает клетка ржи». Эти клетки образуют типичные для ржи 12 хромосом. Форма и число хромосом обусловлены предшествующей историей клетки и зависят от условий их образования. Хромосомы не представляют собой в этом отношении чего-нибудь исключительного. Теми же качествами обладают и другие форменные элементы клетки: миофибриллы, оболочки и т. д.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 