Бурые водоросли отличаются наличием в их хлоропластах, кроме хлорофилла, ряда пигментов красного и бурого цвета. Некоторые из этих пигментов встречаются и в зеленых растениях (например, каротин, ксантофилл); характерным для бурых водорослей является фукоксантин, не встречающийся у других групп растений. Окраска бурых водорослей зависит от количественного соотношения указанных пигментов и колеблется от желтовато-зеленой до темно-бурой. Фотосинтезирующие пигменты связаны с хроматофорами однообразной формы в виде овальных пластинок или зерен. Из других особенностей надо отметить очень распространенное среди бурых водорослей строение оболочки, внутренние слои которой состоят из целлюлозы, а наружные — часто пектиновые, сильно разбухающие, ослизняющиеся в воде. Протопласт диференцирован на ядро и цитоплазму.
Продуктом фотосинтеза является не крахмал, а разные сахара.
Размеры тела весьма разнообразны, как и степень расчлененности; некоторые представители бурых водорослей достигают огромных размеров — до 100 м и больше.
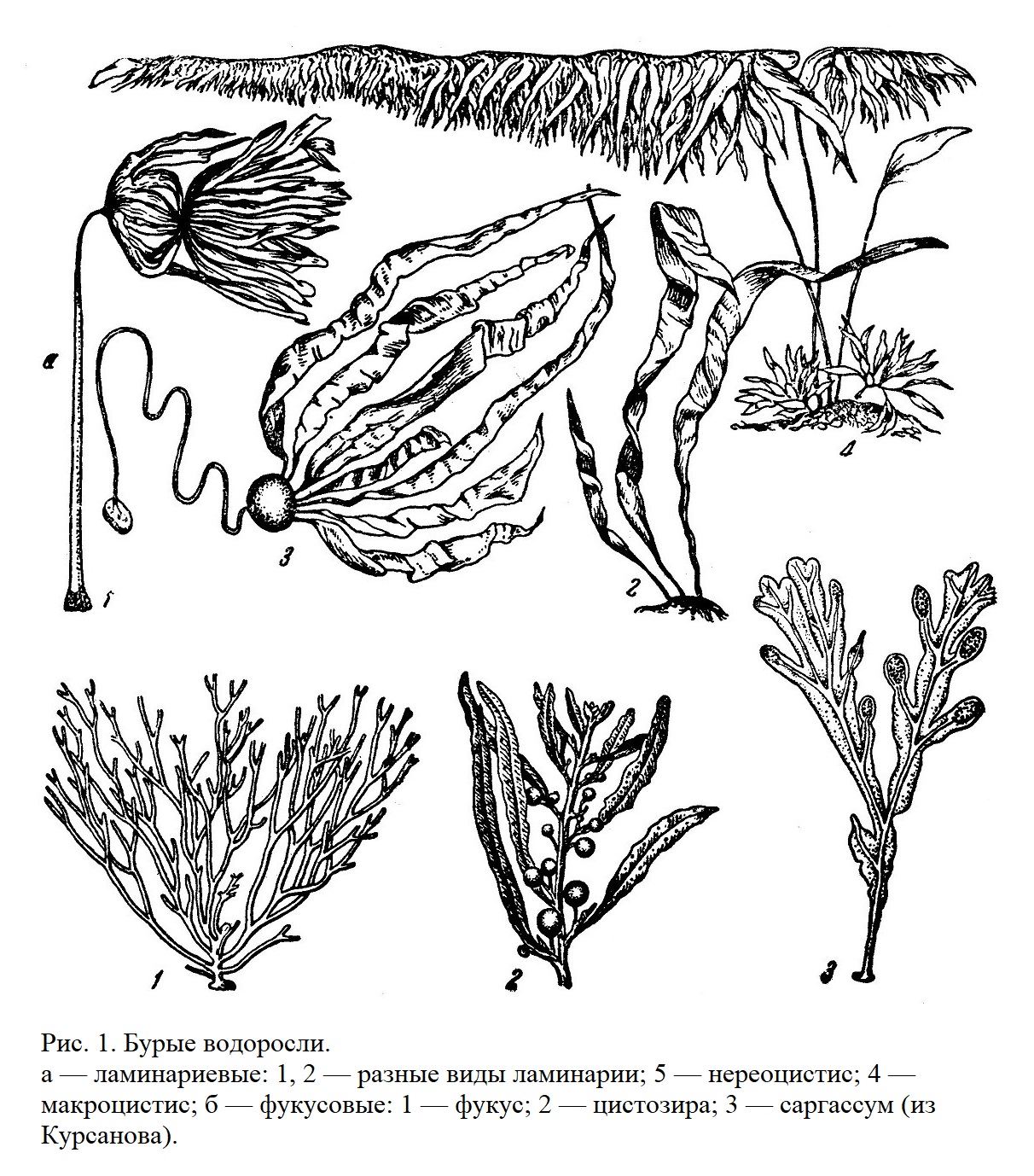
Подавляющее большинство бурых водорослей — обитатели моря притом организмы прикрепленные; они живут большей частью на незначительной глубине. Степень расчленения тела очень велика (рис. 1).
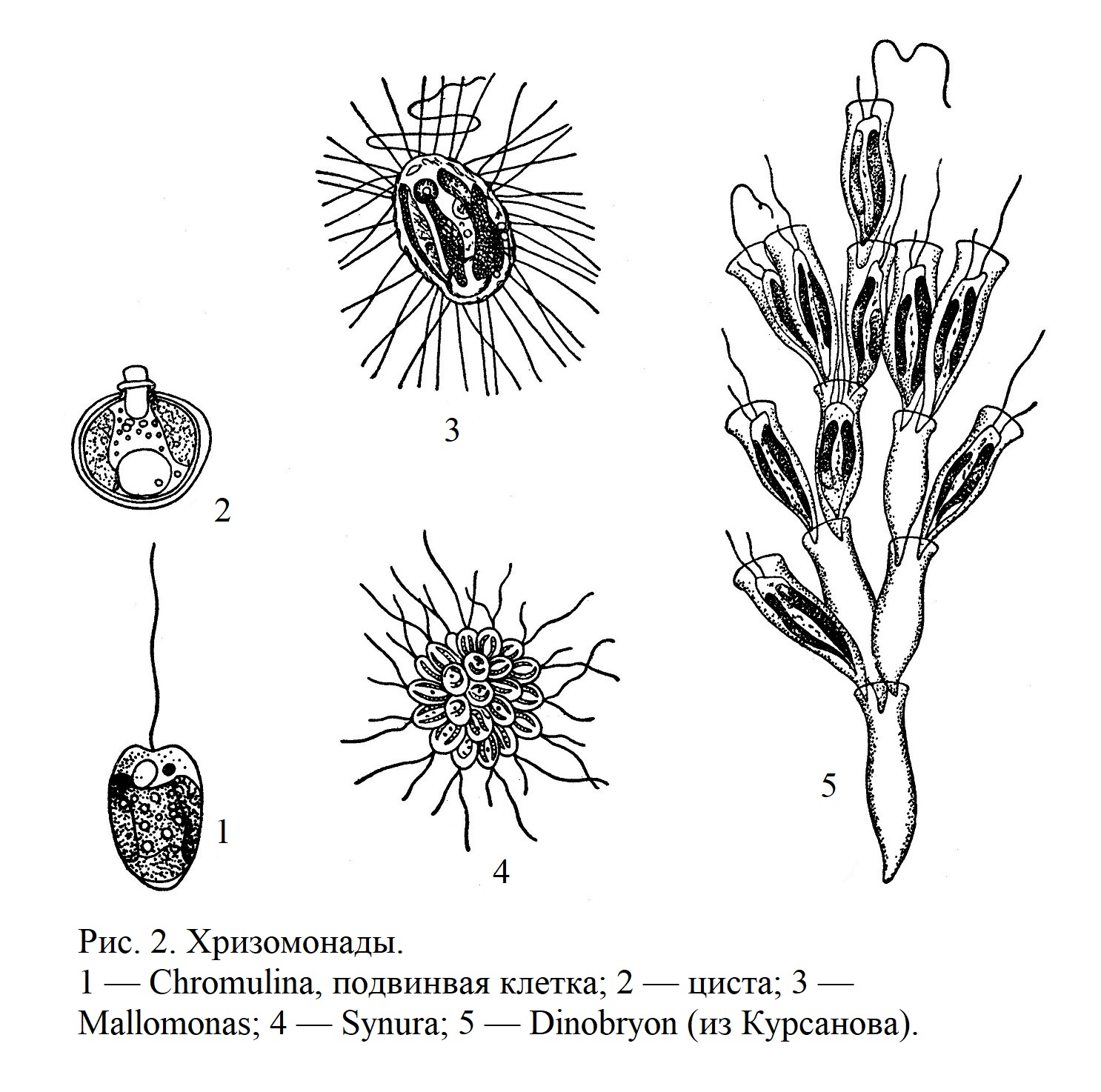
Одноклеточных представителей среди бурых водорослей нет. Наличие жгутиконосных стадий в цикле развития свидетельствует о жгутиконосных предках; однако такие непосредственно переходные формы, какие мы видели у зеленых водорослей, здесь отсутствуют. Возможно, что предками бурых водорослей были организмы, сходные с некоторыми современными жгутиковыми, также окрашенными в бурый цвет,—хризомонадами (рис. 2).
Остатки первых бурых водорослей известны из очень древних пластов, относящихся к ранним периодам палеозойской эры.
Размножение происходит как половым путем (разные формы от изо-гамии до типично выраженной оогамии), так и бесполым (зооспорообразование) (рис. 3 и 5).
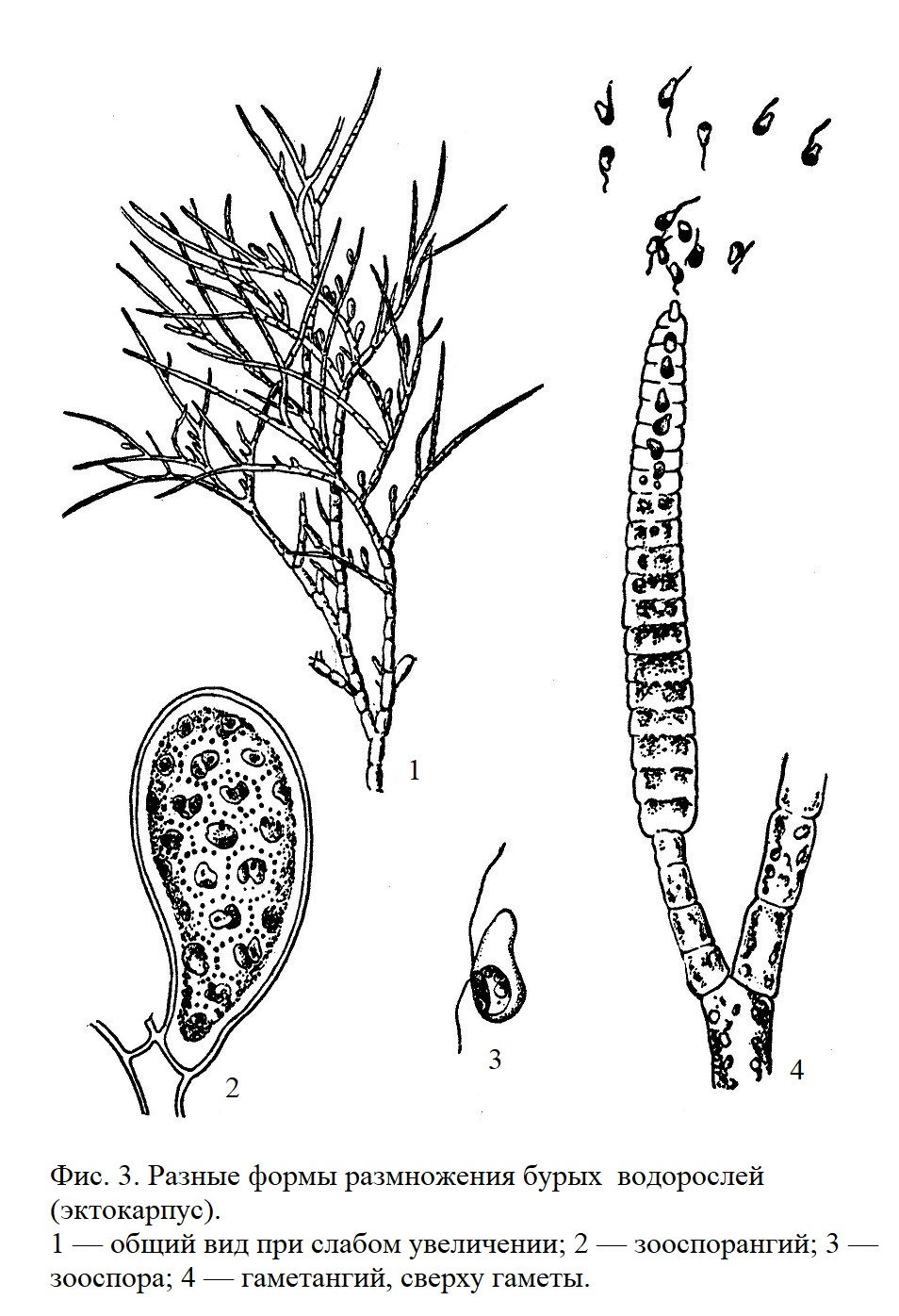
У многих видов хорошо выражено чередование поколений, наблюдаемое и у некоторых зеленых водорослей. Это явление представляет собой смену в цикле развития двух качественно различных форм, называемых половым и бесполым поколениями. Различие между этими поколениями хорошо выражено у некоторых бурых водорослей и морфологически.
В качестве примера такого чередования поколений рассмотрим крупную морскую водоросль ламинарию (рис. 4).

Тело водоросли расчленено на стеблевидную часть, достигающую почти 0,5 м в длину, и листовидную рассеченную пластинку, достигающую 1 — 3 м в длину. Пластинка эта ежегодно сбрасывается и заменяется новой (рис. 4). У основания тела имеются разветвленные бесцветные ризоиды, плотно прикрепляющиеся к подводным камням. При наступлении периода размножения (у европейских видов это связано с осенним похолоданием) описанная водоросль продуцирует только зооспоры; следовательно, она представляет собой организм, размножающийся бесполым путем — бесполое поколение, спорофит. Долгое время у ламинарии неизвестно было другого способа размножениия, а судьба зооспор не была прослежена.
Только в текущем столетии выяснено, что зооспоры, прорастая, дают микроскопически малое растеньице, размножающееся половым путем. Это половое поколение — гаметофит, разные особи которого продуцируют гаметы разных полов. Таким образом, гаметофит ламинарии является раздельнополым микроскопическим растеньицем (рис. 4). Одни растеньица состоят из коротких не ветвистых нитей, на которых развиваются округлые яйцеклетки, находящиеся по одной в так называемых оогониях — крупных вздутых клетках нити. Это — женские растеньица, женский гаметофит. Мужской гаметофит представляет собой более длинные, слегка ветвящиеся нити, на которых образуются мелкие вздутия — антеридии — органы, в которых развивается по одному маленькому, подвижному, снабженному жгутиками, сперматозоиду. Созревшие сперматозоиды выплывают из антеридия, подплывают к яйцеклетке, происходит оплодотворение — слияние сперматозоида с яйцеклеткой.
Развитие гаметофита происходит медленно, в течение всей зимы. Образование половых органов и половых клеток происходит уже весной.
Возникшая после оплодотворения зигота прорастает тотчас, без периода покоя, и дает новое поколение, резко отличное от гаметофита; из зиготы развивается уже описанный спорофит — большая, сложно расчлененная водоросль. Таким образом, морфологически отличные друг от друга поколения различаются также и биологически: развитие одного ведет к образованию спор, другого — к образованию гамет.
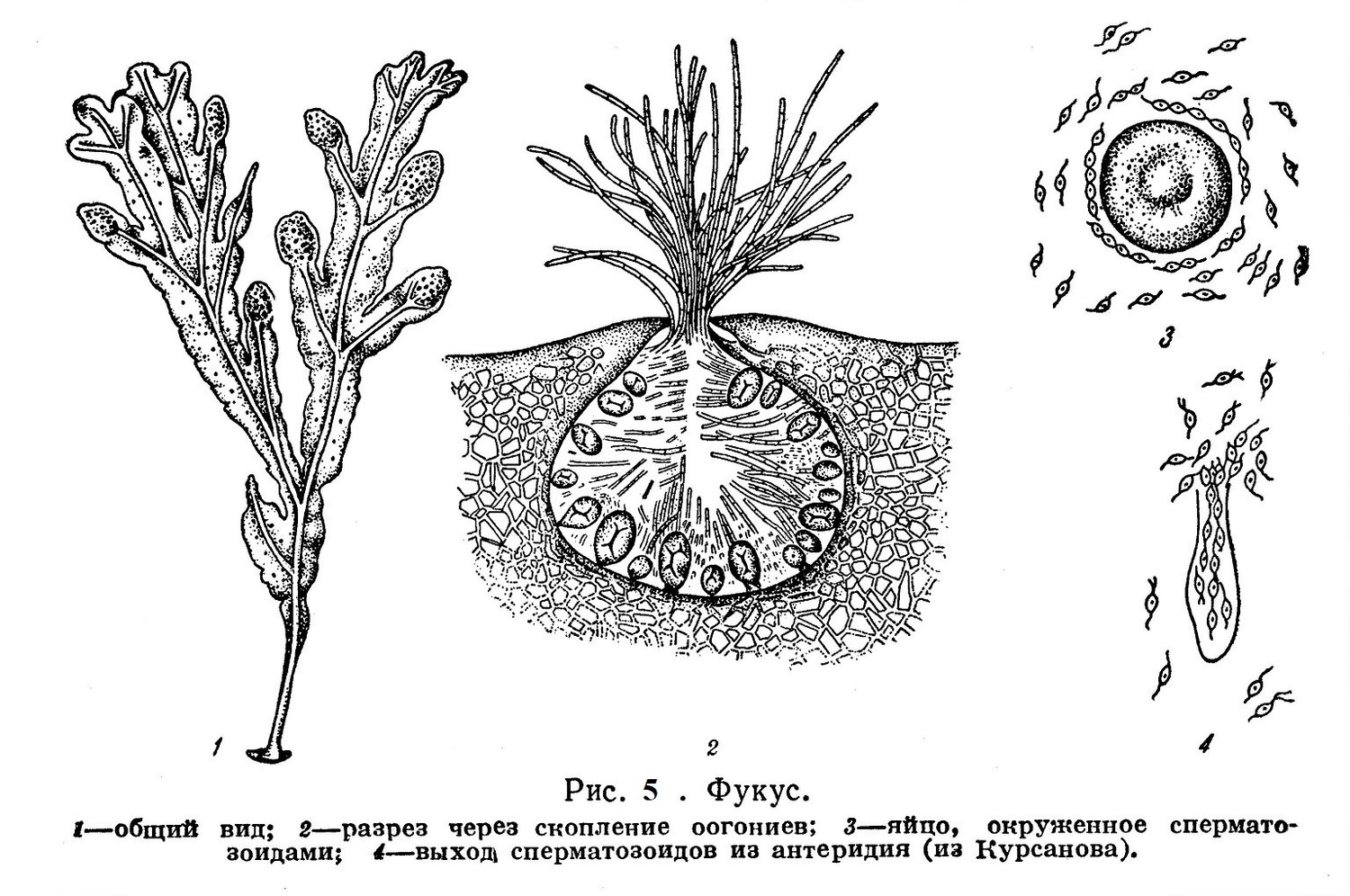
Мужские гаметы бурых водорослей снабжены жгутиками, имеют хроматофор и напоминают зооспоры. На рис. 5 изображен процесс оплодотворения у бурой водоросли фукус.
Бурые водоросли живут и в теплых, тропических частях океанов. Немногие из этих водорослей поселяются на значительных глубинах. Глубоководные бурые водоросли, прикрепляясь к придонным камням, подымают высоко к поверхности моря листовидные части своего слоевища; такому вертикальному положению способствуют воздухоносные пузыри, возникающие на их слоевище.
Сюда относятся такие гиганты, как макроцистис и др., в изобилии произрастающие в Саргассовом море (в тропической части Атлантического океана) (рис. 1).
Относительно значения бурых водорослей можно повторить многое из того, что сказано о значении водорослей вообще. Как и все водоросли, они являются источником питания морских животных.
Многие из них употребляются в пищу человеком, в частности, некоторые виды ламинарий и др., известные под названием морской капусты. Многие служат кормом скоту. Имея в составе своего тела большое количество калия, эти водоросли являются ценным калийным удобрением и сырьем для добычи калия.
Некоторые бурые водоросли содержат, как и многие красные, большое количество йода и являются источником его добывания. Отсюда их диететическое и лекарственное значение при применении в пищу.
Из бурых водорослей, как и из красных, добывается особое желеподобное вещество агар-агар, смесь разных полисахаридов; он широко применяется в микробиологической методике, а также в кондитерском производстве Из бурых водорослей добывают и другие клеевые вещества, употребляемые в текстильном и других производствах.
Стеблевидные части слоевища некоторых бурых водорослей (ламинария) употребляются иногда в медицинской практике с целью расширения отверстий при оперативном вмешательстве, это основано на способности оболочек ослизняться и сильно набухать.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 