Ранее мы остановились на описании той стадии развития зачатка сосудистой крови, когда в нем образовались первичные кровяные клетки, или первичные гемоцитобласты. Последние представляют довольно крупные клетки округлой формы с базофильной цитоплазмой и ядром, в котором хорошо обнаруживаются крупные глыбки хроматина. Эти клетки способны к амебоидным движениям и представляют такую же клеточную генерацию, как уже упоминавшиеся выше первые блуждающие клетки в мезенхиме.
В сосудах желточного мешка первичные кровяные клетки усиленно размножаются митозами, и громадное их большинство одновременно преобразуется в первичные эритробласты (рис. 139). Это превращение совершается так, что в цитоплазме первичных кровяных клеток (гемоцитобластов) накапливается гемоглобин, и цитоплазма постепенно приобретает все более и более оксифильный характер.
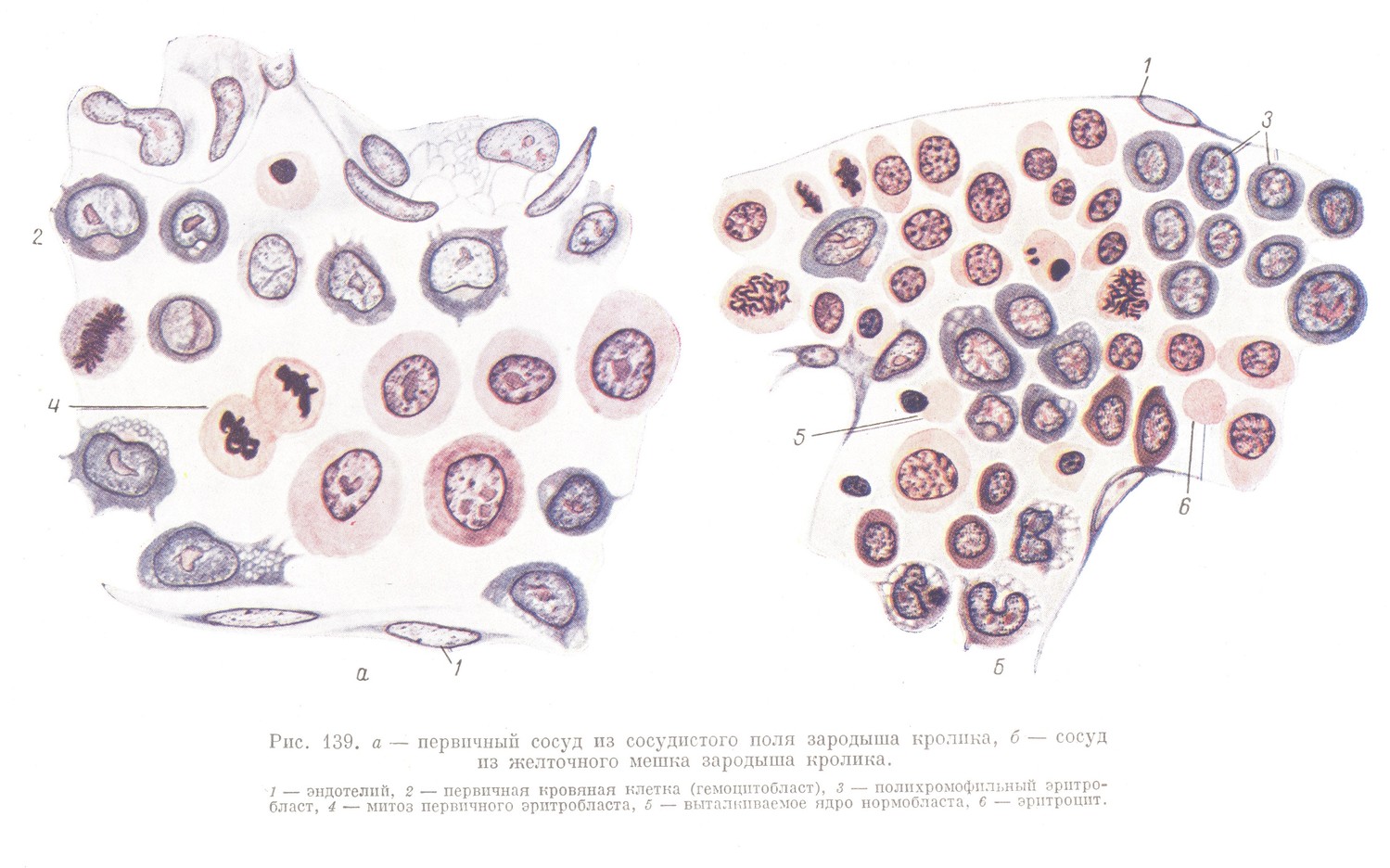
Первичные эритробласты продолжают митотически размножаться, вследствие чего количество их в циркулирующей крови увеличивается (рис. 139, а).
Одновременно с размножением нарастают изменения регрессивного характера, выражающиеся в увеличении количества гемоглобина и в уплотнении и пикнотизации ядра. В дальнейшем эти изменения заходят столь далеко, что первичные эритробласты утрачивают способность к размножению и превращаются в первичные эритроциты, которые позже могут стать безъядерными в связи с выталкиванием из своего тела переставшего функционировать пикнотического ядра. Выброшенные ядра и утратившие жизнеспособность эритроциты поглощаются фагоцитами, происходящими из сосудистого эндотелия.
Новообразование первичных эритробластов из первичных кровяных клеток прекращается на сравнительно ранних стадиях развития. Поэтому после того, как все первичные эритробласты в процессе диференцировки превратятся в первичные эритроциты, а эти последние в дальнейшем дегенерируют, все первичные гемоглобинные элементы окажутся исчезнувшими из организма. Это наступает во второй половине утробной жизни.
На ранних стадиях развития освобождающиеся из мезенхимного синцития блуждающие клетки мало отличаются от первичных кровяных клеток и могут дать начало таким же эритробластам, как и в сосудистых зачатках. Большинство первичных блуждающих клеток мезенхимы утрачивает свою резкую базофилию, цитоплазма их становится ячеистой, и они преобразуются в тех гистиоцитов, которые были описаны выше при рассмотрении соединительной ткани.
Не все первичные кровяные клетки диференцируются в первичные эритробласты. Часть гемоцитобластов остается в малодиференцированном состоянии и дает начало поколениям малодиференцированных блуждающих элементов, расселяющихся по всей соединительной ткани и крови. Эти клетки становятся родоначальниками всех дальнейших кроветворных процессов, почему они и получили название гемоцитобластов.
Из таких гемоцитобластов еще в сосудах желточного поля начинается образование вторичных эритроцитов. Последнее происходит также путем диференцировки, которая совершается параллельно с размножением, прекращающимся лишь на последнем этапе превращения эритробластов в эритроциты. Образование вторичных эритроцитов по сравнению с первичными эритробластами идет более медленно и на некотором отрезке времени совершается одновременно с первичным эритропоэзом. В процессе развития из гемоцитобласта вторичные эритроциты проходят стадию полихроматофильной эритробласта с небольшим содержанием гемоглобина и в дальнейшем длительную стадию эритробласта с постепенным нарастанием содержания гемоглобина и прогрессирующей пикнотизацией ядра. Развивающиеся вслед за тем элементы, накапливающие в себе еще больше гемоглобина, все же сохраняют способность к размножению и называются нормобластами.
На последней стадии развития зрелый нормобласт утрачивает способность к делению; ядро выталкивается и эритроцит становится безъядерным (рис. 139, б). Такие вторичные эритроциты мало отличаются от эритроцитов крови взрослого животного и по сравнению с первичными эритроцитами имеют более правильную форму и несколько меньшую величину (в особенности это относится к первичным эритробластам).
После редукции желточного мешка местом образования вторичных эритроцитов (эритропоэза) становится печень, где этот процесс совершается уже не внутри сосудов, а вне их, экстраваскулярно; после образования первых костей окончательным местом образования эритроцитов становится костный мозг, выполняющий функцию кроветворения и во взрослом организме. После перемещения центра эритропоэза в костный мозг кроветворные очаги печени редуцируются. Обычно это происходит еще до рождения.
Источником кроветворения и в печени, и в костном мозге является все тот же гемоцитобласт (рис. 140). Гемоцитобласты могут иметь местное происхождение (из мезенхимы, окружающей сосуды, и даже из эндотелия), а также происходить из принесенных током крови первичных кровяных клеток, образовавшихся где-либо в другом месте.
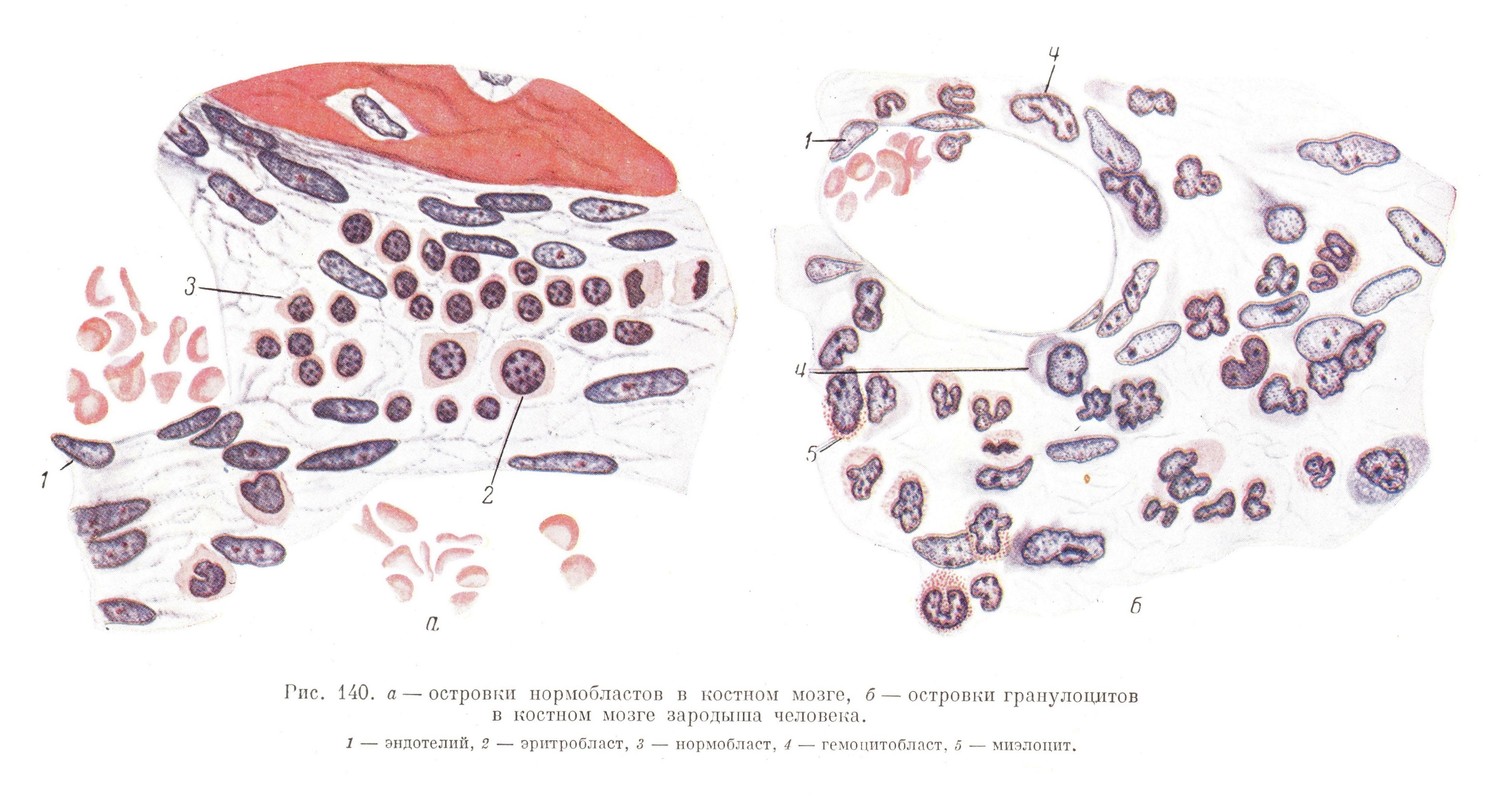
И в печени, и в костном мозге, кроме эритроцитов, начинают развиваться и зернистые лейкоциты (главным образом специальные и эозинофильные), развитие которых идет вначале таким же укороченным путем, как и развитие первичных эритробластов. В цитоплазме гемоцитобласта, которая становится более светлой и менее базофильной, появляется специфическая зернистость, после чего ядро приобретает неправильную форму и гранулоцит получается сразу, без промежуточной стадии миэлоцита (рис. 140).
Кроме гранулоцитов, в кроветворной ткани печени и костного мозга образуется еще один клеточный элемент, происхождение которого еще не вполне выяснено; это — гигантские клетки, или мегакариоциты.
Остановимся на кроветворении в костном мозге, строение которого будет подробно изложено в дальнейшей нашей части сайта.
Кроветворные участки костного мозга состоят из миэлоидной ткани, которая развивается из мезенхимы в процессе образования кости.
Мезенхимный синцитий с проходящими в нем капиллярами на первых стадиях развития костного мозга образует его строму. В дальнейшем большая часть мезенхимного синцития превращается в ретикулярную ткань с нежным ретикулиновым остовом, тогда как другая, незначительная его часть (в окружности более крупных сосудов) образует рыхлую соединительную ткань. Ретикулярные элементы стоят в непосредственной цитоплазматической связи с эндотелием капилляров, который обладает свойствами ретикулярной ткани. Часть клеток ретикулярной ткани, накапливая в цитоплазме жир, превращается в жировые клетки. В таком виде строма костного мозга находится во взрослом организме.
В начале развития костный мозг никаких кроветворных элементов, кроме редко разбросанных гемоцитобластов как местного происхождения, так и пришлых, не содержит. Эти гемоцитобласты, так же как и в печени, становятся родоначальниками кроветворных процессов костного мозга, Продуцирующего в нормальных условиях эритроциты и гранулоциты.
В процессе развития костного мозга раньше всего начинается эритропоэз, который происходит по той же схеме, что и в печени, т. е. претерпевает стадии проэритробластов, эритробластов, нормобластов и эритроцитов (рис. 140).
Все: клеточные формы, до нормобластов включительно, способны к митотическому размножению, в процессе которого и совершается их прогрессирующая диференцировка по схеме, приведенной ниже. Зрелые нормобласты утрачивают способность к делению, теряют ядро и становятся эритроцитами.
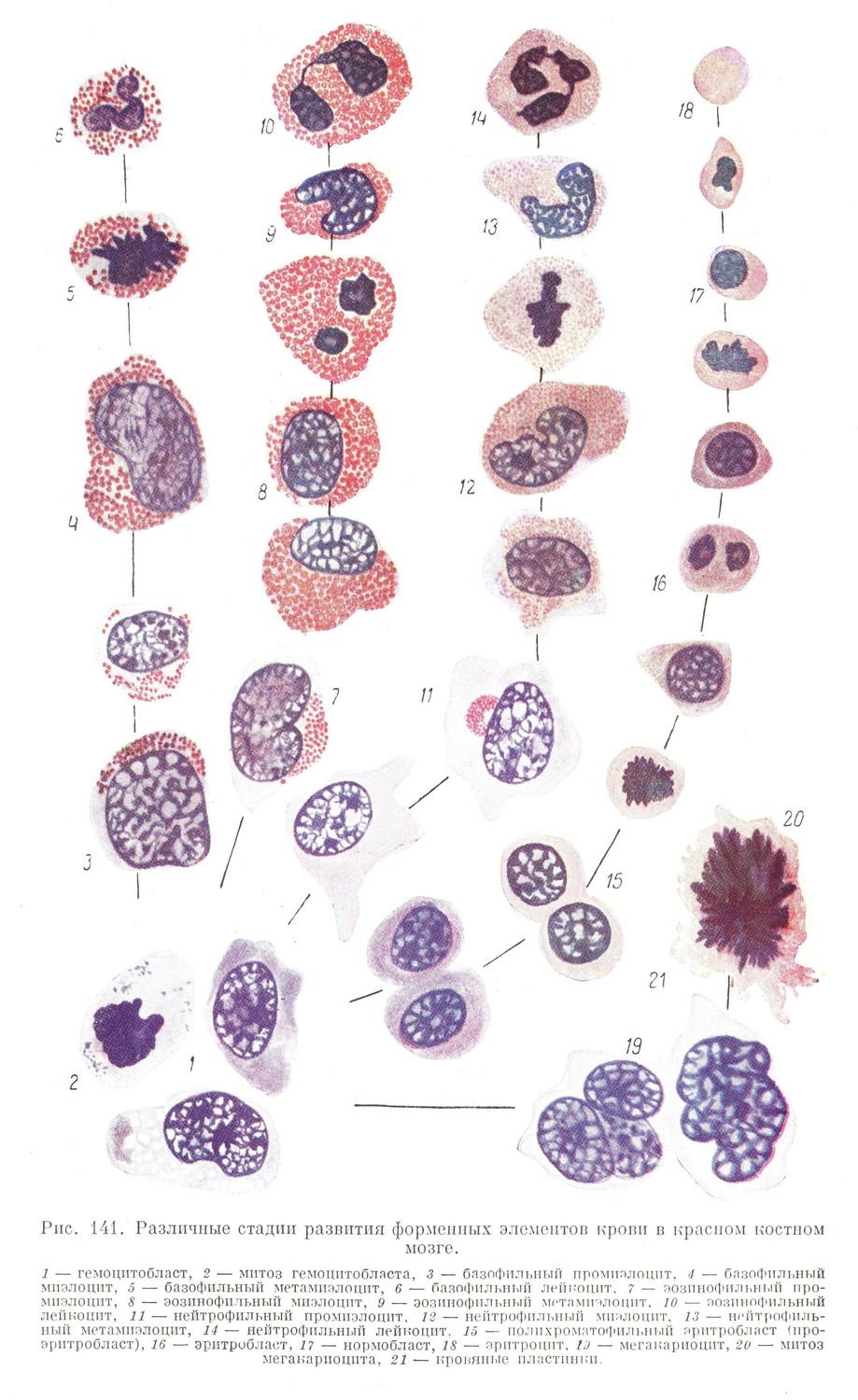
В костном мозге взрослого в нормальных условиях главная масса кроветворных клеток эритробластического ряда представлена уже значительно более диференцированными эритробластами и даже нормобластами, которые непрерывно размножаются и создают основное вещество для образования эритроцитов (рис. 141). Таким образом, в нормальных условиях в костном мозге содержится огромное количество почти готового резервного материала для пополнения убыли в эритроцитах, из которого может быстро восстанавливаться нормальное число эритроцитов крови даже при значительных кровопотерях.
В этих случаях на пополнение убыли эритроцитов в сосудистой крови растрачивается большая или меньшая масса эритробластов, и тогда начинается восстановление их запаса путем размножения гемоцитобластов и даже их новообразование из ретикулярного синцития с последующей диференцировкой, проходящей через стадию полихроматофильных эритробластов.
Относительно исчезновения ядра из нормобластов (при превращении их в эритроциты в процессе кроветворения во взрослом организме) существуют два мнения. Одни исследователи считают, что ядро исчезает благодаря каким-то аутолитическим процессам, растворяясь в теле самого нормобласта; другие полагают, что ядро из его тела выталкивается и пожирается макрофагами, происходящими из ретикулярного синцития. Более вероятно, что существуют и тот и другой способы редукции ядра.
Подобным образом совершается и образование белых элементов — гранулоцитов (рис. 141). Первые гранулоциты в костном мозге, дак же как и в печени, образуются, как это было отмечено, укороченным путем и представляют формы не вполне зрелые, иногда еще способные к размножению. На более поздних стадиях развития и во взрослом организме гранулопоэз протекает более медленным темпом. Источником новообразования являются те же гемоцитобласты, которые, размножаясь митотическим путем, начинают диференцироваться одновременно в трех различных направлениях и продуцируют миэлоциты трех видов: специальные, эозинофильные и базофильные.
Специальные миэлоциты вначале (промиэлоциты) представляются клетками большей, чем гемоцитобласты, величины с базофильной, но более бледной цитоплазмой, лишенной зернистости, и с округлым светлым ядром, в котором хорошо видны ядрышки. Позднее в цитоплазме начинает появляться специфическая зернистость (у человека несколько более оксифильная, чем в диференцированном нейтрофиле), которая все более и болое заполняет цитоплазму.
У более старых миэлоцитов (метамиэлоцихов) ядро постепенно становится плотнее и приобретает полиморфный вид. Наконец, миэлоцит утрачивает способность к делению и превращается в зрелый специальный гранулоцит.
В нормальном костном мозге содержатся главным образом метамиэлоциты и готовые молодые лейкоциты. Таким образом, и в этом случае, так же как и в случае эритробластов, в костном .мозге содержится почти вполне готовый резерв, из которого быстро может пополниться убыль специальных лейкоцитов.
Аналогичным путем идет образование и эозинофилов, и базофилов. Появляющаяся в эозинофильных миэлоцитах молодая зернистость обладает некоторым сродством к основным краскам и только при созревании приобретает резко выраженную оксифильность.
В существовании базофильных миэлоцитов некоторые авторы сомневаются. Однако для млекопитающих и человека факт существования специального ствола миэлоцитов, дающего начало базофильным гранулоцитам, следует считать доказанным.
Из гемоцитобластов костного мозга происходят и гигантские клетки или мегакариоциты. Они образуются путем гипертрофии ядра, амитотического его перешнуровывания, наступления многополюсных митозов без деления клеточного тела и т. д. (рис. 141). В результате получаются гигантские (по сравнению с другими элементами миэлоидной ткани) клетки, с мутной, содержащей азурофильные включения, цитоплазмой, неправильным ядром (или ядрами) и множественным центром.
К размножению эти клетки, по-видимому, уже не способны. От их тела постоянно отделяются частицы цитоплазмы с азурофильными зернами. Этот факт дает повод считать мегакариоциты источником образования кровяных пластинок.
Наряду с этими специфическими для костного мозга кроветворными элементами, в нем в нормальных условиях всегда содержится некоторое количество лимфоцитов, моноцитов и продуктов их превращения — плазматических клеток. Большинство авторов считает лимфоцитов, обнаруживаемых в костном мозге, элементами пришлыми, образующимися в других местах.
Лимфоциты появляются в организме зародыша на более поздних стадиях, чем эритроциты, — так же как и зернистые лейкоциты. Только у млекопитающих и человека имеются локализованные участки лимфоидного кроветворения в виде лимфатических узлов (рис. 142). У остальных позвоночных лимфоциты образуются в диффузных очагах, рассеянных в соединительной ткани, богатой недиференцированными элементами (например в слизистых оболочках).
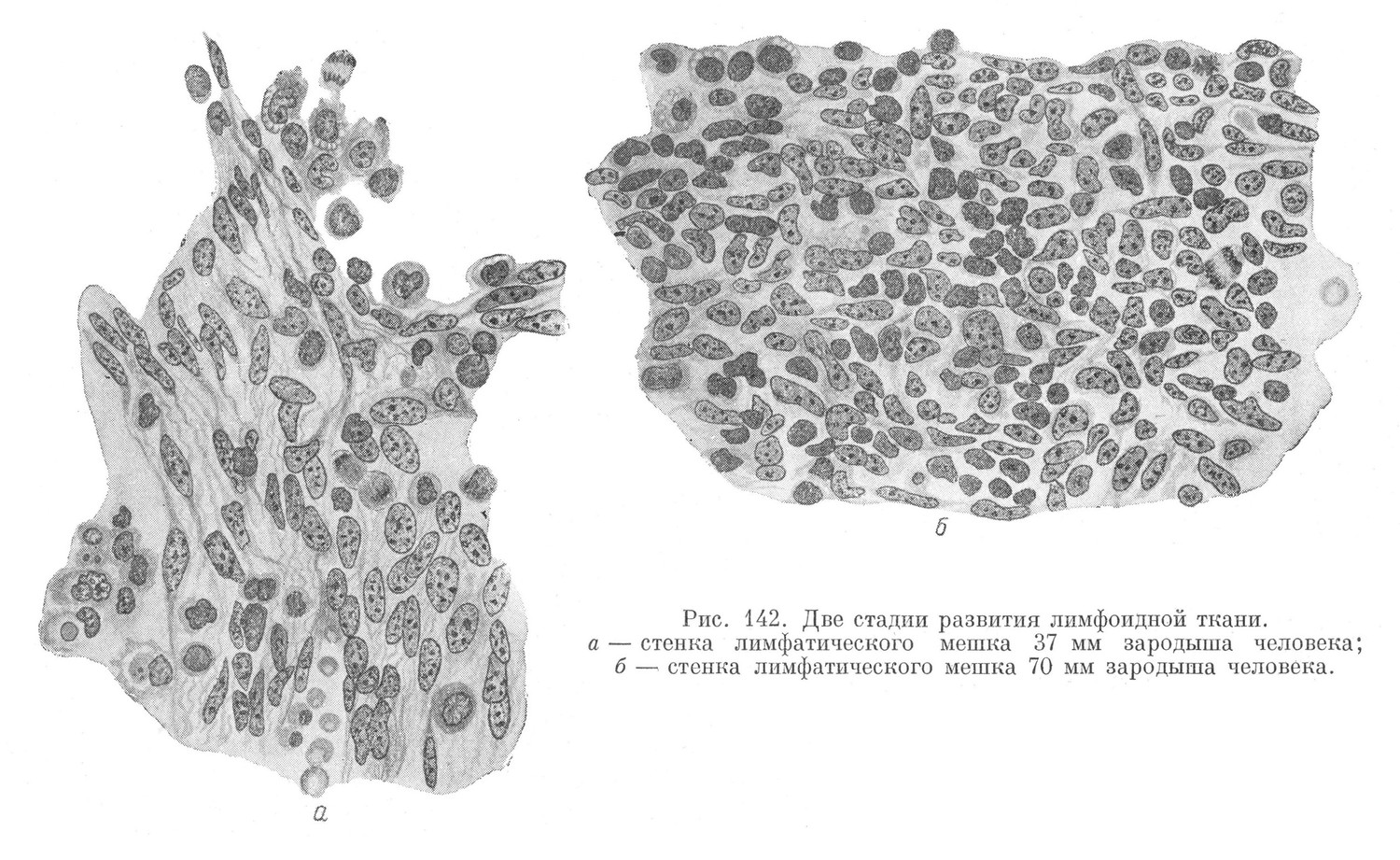
Первые зачатки лимфатических узлов появляются, в области шейных лимфатических мешков. Лимфатические узлы задних областей тела закладываются позже.
В мезенхиме стенки лимфатического мешка начинают образовываться различные свободные элементы непосредственно из мезенхимного синцития. Основными элементами являются, во-первых, клетки типа большого лимфоцита или гемоцитобласта, не отличимые от гемоцитобластов других кроветворных участков, и, во-вторых, малые и средние лимфоциты, которые отличаются от гемоцитобластов меньшими размерами и более темным ядром. Малые лимфоциты вначале образуются непосредственно из мезенхимных клеток, т. е. таким же укороченным путем, как первые эритробласты и первые гранулоциты. В дальнейшем прямое превращение мезенхимных клеток в малые лимфоциты постепенно прекращается, и главным источником для развития последних становятся гемоцитобласты.
Образование малых лимфоцитов из гемоцитобластов происходит ужа более длинным путем, так же как и в лимфоидных органах взрослого организма, т. е. проходит стадию среднего лимфоцита. Мезенхимный синцитий в области развивающегося узла постепенно преобразуется в ретикулярную ткань, в петлях которой и накапливаются различные свободные элементы. Кроме гемоцитобластов и лимфоцитов, здесь постоянно встречаются гистиоциты, или макрофаги, образующиеся и из гемоцитобластов и непосредственно из ретикулярной ткани.
Следует отметить, что на ранних стадиях развития в зачатках лимфатических узлов наблюдается присутствие миэлоидных элементов — эритробластов и гранулоцитов (с укороченным развитием), мегакариоцитов и даже миулоцитов. Однако размножение этих форм быстро прекращается и подавляется образованием лимфоцитов, которые и составляют главную массу свободных элементов лимфоидной ткани.
Образование лимфоцитов происходит по следующей схеме. Исходной формой является гемоцитобласт, который, размножаясь, дает поколение лимфоцитов средних размеров; продуктами размножения последних являются уже малые лимфоциты. В лимфатических узлах, как и в кровяном русле, малые лимфоциты почти не делятся. Таким образом, на основании изучения только лимфоидного кроветворения мы вправе сделать вывод о том, что малый лимфоцит является законченной, неспособной к размножению диференцированной формой, аналогичной гранулоцитам.
Однако ряд фактов, относящихся к поведению малых лимфоцитов при воспалительных процессах и в культурах in vitro, указывает на их способность к прогрессивным превращениям, а также и к делению. Таким образом, при характеристике малого лимфоцита возникают значительные противоречия, для примирения которых возможен только один выход, а именно — допущение, что лимфоциты (в смысле их диференцировки) являются такой же неоднородной массой, как, например, фибробласты. Следовательно, часть из них представляет формы малодиференцировацные и способные к прогрессивным изменениям (при воспалении и в культурах), а другая часть, напротив, достигает такой степени диференцировки, что утрачивает способность и к делению и к дальнейшей диференцировке. В кровяном русле нормальной крови попадаются и те и другие. Отсюда и возникают противоречия. В лимфоидной ткани, как правило, встречаются и плазматические клетки, являющиеся продуктами старческой инволюции лимфоцитов (и моноцитов).
Из предыдущего изложения можно убедиться в том, как постепенно диференцируются в процессе развития из общего мезенхимного зачатка различные производные последнего. Наряду с диференцированными элементами — эритроцитами, фиброцитами, гранулоцитами и (может быть, частично) малыми лимфоцитами, здесь имеются и малодиференцированные элементы — камбиальные клетки и гемоцитобласты, а также малоспециализированные, способные к дальнейшему, далеко идущему развитию ретикуло-эндотелиальные элементы и часть лимфоцитов. К подобным же малоспециализированным элементам следует отнести и моноциты, источником происхождения которых являются, главным образом, ретикулярные клетки и гистиоциты.
Хотя все элементы трофической системы и происходят из общего корня, каковым являются эквипотенциальные мезенхимная клетка и гемоцитобласт, однако образование различных элементов крови и соединительной ткани сосредоточено в определенных местах.
У человека и млекопитающих таким очагом для образования эритроцитов и гранулоцитов является красный костный мозг, для лимфоцитов — лимфатические узлы и другие скопления лимфоидной ткани. Это обстоятельство имеет важное клиническое значение. При тех заболеваниях крови, которые называются лейкемиями, картина периферической крови обычно указывает на нарушения функций либо в миэлоидной, либо в лимфоидной системах. В первом случае состав эритроцитов и гранулоцитов (миэлоидная лейкемия) бывает ненормальным, во втором — изменяется состав малых лимфоцитов (лимфоидная лейкемия).
В последнее время установлен еще один вид лейкемического изменения крови, выражающийся в ненормальном повышении числа моноцитов, что трактуется как выражение патологических изменений в рети- куло-эндотелиальной системе, которая является главным источником их образования.
Все эти клинические данные послужили поводом для обоснования широко распространенной в настоящее время триалистической теории кроветворения, согласно которой родоначальные формы миэлоидных элементов (эритроцитов и гранулоцитов), лимфоцитов и моноцитов представляют различные не переходящие одна в другую формы.
Однако такая точка зрения мало согласуется с изложенными выше фактами эмбрионального кроветворения, с тем, что в каждом очаге вначале имеются все элементы, и только впоследствии процесс образования некоторых элементов подавляется и развиваются преимущественно элементы определенного характера: в одних случаях — миэлоидные, в других — лимфоидные. При изменении условий часто происходит преобразование лимфоидных очагов в миэлоидные (миэлоидная метаплазия лимфатических узлов) и, наоборот, — миэлоидных органов в лимфоидные (лимфоидная метаплазия костного мозга). Кроме того, иногда на различных участках соединительной ткани могут возникать спонтанные очаги то миэлоидного, то лимфоидного кроветворения.
Все эти факты дали повод к построению так называемой унитарной теории кроветворения, согласно которой все кровяные элементы происходят из единой, общей для всех, родоначальной клетки — гемоцитобласта. При попытках объяснить патологические картины крови эта теория, однако, мало убедительна.
Кроме унитарной и триалистической теории, существует еще дуалистическая, согласно которой кровяные клетки происходят из двух элементов, а также различные полифилетические теории, которые предполагают наличие более трех родоначальных форм.
Многочисленность противоречивых теорий указывает, несомненно, на принципиально неправильный подход к постановке вопроса о кроветворении. Основная ошибка, по нашему мнению, состоит в том, что авторы множественных родоначальных теорий пытаются приписать им абсолютную детерминированность, а авторы монистических воззрений — абсолютную лабильность (неустойчивость). Между тем, весьма вероятно, что для камбиальных элементов тканей внутренней среды не существует ни абсолютной детерминированности, ни абсолютной лабильности.
Имеется много данных, подтверждающих, что в известных условиях кроветворный камбий детерминирован лабильно. Лабильная детерминированность обусловливает те факты, которые лежат в основе всех теорий о множественных родоначальных формах кроветворения. При более резких вмешательствах и более значительных общих или местных изменениях эта лабильная детерминированность нарушается, и тогда создаются условия для различных перестроек в кроветворении, которые дали повод к обоснованию монистических воззрений.
У прочих позвоночных (кроме млекопитающих и птиц) и эмбриональное, и окончательное кроветворение изучено значительно хуже. Из того, что известно по этому вопросу, можно, однако, заключить, что в общем оно совершается так же, как и у млекопитающих, с той лишь разницей, что локализация (окончательная) очагов кроветворения бывает различная и часто иная, чем у млекопитающих.
Прежде всего образование лимфоцитов (за исключением некоторых птиц), как это уже было упомянуто, происходит не в локализованных очагах, а повсеместно в соединительной ткани. Далее образование эритроцитов происходит всегда внутри сосудов (интраваскулярно), а не вне их. Этот васкулярный эритропоэз локализуется, в конце концов, то в костном мозге (птицы, рептилии, бесхвостые амфибии), то в селезенке (рыбы, хвостатые амфибии).
Образование гранулоцитов совершается экстраваскулярно также в различных местах: в костном мозге (птицы, рептилии и бесхвостые амфибии), в почке, в кишечнике (рыбы), в печени (хвостатые амфибии) и т. д. Тромбоциты образуются там же, где развиваются эритроциты.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 