Все отправления печени можно разделить на 4 основные функции: 1) экскреторную, 2) трофическую, 3) защитную и 4) кроветворную (в эмбриональном периоде).
Экскреторная функция печени заключается в выработке желчи и мочевины. И та и другая вырабатываются клетками печеночной паренхимы из веществ, приносимых кровью. Желчные пигменты образуются при этом из содержащих железо продуктов разрушения гемоглобина, приносимых в печеночную кровь, главным образом, селезеночной веной. Желчь выделяется через выводные протоки печени в кишку и выбрасывается вместе с калом наружу. Таким образом, желчь является отбросом организма, экскретом. Но одновременно она используется в пищеварительном процессе как вещество, нейтрализующее кислый желудочный сок, и как вещество, способствующее омылению и резорбции жиров. Хотя желчь и не содержит ферментов, однако нормального усвоения жиров без нее не происходит.
Мочевина в печени образуется из приносимых кровью азотистых продуктов обмена, обладающих обычно ядовитыми свойствами. В печеночной паренхиме эти вещества преобразуются в менее ядовитую мочевину, которая и выделяется снова в кровь. Таким образом, кровь, выходящая из печени, содержит значительно большее количество мочевины, чем кровь воротной вены. Образовавшаяся в печени мочевина выделяется почками.
Трофическая функция печени у высших животных и у человека заключается, главным образом, в том, что всасываемые в кишке из пищевой кашицы в кровь углеводы (глюкоза), попадая в печень, в значительной части захватываются печеночными клетками и преобразуются в гликоген (животный крахмал), представляющий нерастворимый полисахарид. Последний, по мере надобности, снова переходит в растворимое состояние и выбрасывается в кровь в виде глюкозы, которая и расходуется органами (главным образом, мышцами). Таким образом, печень служит как бы запасным депо для работающих частей организма (гликогенная функция).
У низших животных в печени накапливается также и резервный жир. У человека и млекопитающих в нормальных условиях жир в печени откладывается в очень незначительных количествах.
Защитная (фагоцитарная) функция печени обусловлена, главным образом, особым строением стенки ее капилляров, являющейся составной частью ретикуло-эндотелиальной системы организма.
Кроветворная функция свойственна печени лишь в эмбриональном периоде и на строении печени во взрослом организме отражается мало.
Из сказанного ясно, что, во-первых, в печени обмен между кровью и железистой паренхимой носит двусторонний характер, 1 во-вторых, печень является органом, выделяющим вырабатываемые вещества преимущественно в кровь, а не во внешнюю среду. Таким образом, печень является железой скорее внутренней, чем внешней секреции, хотя и не вырабатывает никаких специфических веществ (гормонов), как настоящие железы внутренней секреции.
Все эти соображения заставляют предполагать наличие в печени особого строения, облегчающего эту двустороннюю секрецию, что в действительности и обнаруживается.
Для того, чтобы лучше представить строение печени во взрослом организме, необходимо рассмотреть ее в эмбриональном состоянии на такой стадии, когда основные черты ее структуры выступают с особенной отчетливостью.
Печень развивается из выпячивания вентральной стенки двенадцатиперстной кишки, врастающего в вентральную брыжейку. Это выпячивание, имеющее вид плотного тяжа, разделяется на многочисленные веточки, которые, анастомозируя между собой и усиленно разрастаясь, образуют железистую паренхиму печени, постепенно приобретающую трубчатое строение.
В первой половине утробной жизни печень зародыша не обнаруживает подразделения на дольки. Железистая паренхима представлена плотными анастомозирующими клеточными тяжами и напоминает своим строением губку. Между эпителиальными тяжами, плотно прилегая к ним своими стенками и выполняя все свободные промежутки, располагается сеть широких венозных капилляров (на рисунке светлые), происходящих из желточной вены, являющейся родоначальницей воротной вены.
Таким образом, на этой стадии печень своим строением напоминает типичную железу внутренней секреции. Примером строения таких желез могут служить уже описанные выше островки поджелудочной железы.
В печени зародыша имеется очень большая поверхность соприкосновения эпителиальных клеток со стенками широких капилляров. Отличие от островков заключается лишь в том, что эпителиальные тяжи печеночной паренхимы зародыша являются одновременно и зачатками железистых отделов с внешней секрецией, открывающихся выводными протоками в просвет двенадцатиперстной кишки.
Дальнейшая диференцировка такой однородной во всех своих частях печени происходит во второй половине утробной и в начале внеутробной жизни. Происходит эта диференцировка следующим образом.
По ходу воротного комплекса сосудов внутрь печени врастает соединительная ткань являющаяся прямым продолжением фиброзной капсулы, покрывающей печень снаружи. Эта ткань разделяет печеночную паренхиму на отдельные территории, называемые дольками. Печеночные дольки отличаются от долек других желез тем, что все они приблизительно одинаковые и соответствуют долькам самого низшего порядка прочих желез (где мелкие дольки собираются в более крупные, эти последние в еще более крупные и т. д., так что дольки высшего порядка являются уже макроскопически хорошо заметными образованиями).
Печень макроскопически кажется однородной, ее дольки не превышают размеров 1—1,5 мм и слабо видимы невооруженным глазом. Междольковая соединительная ткань более или менее значительно развита только у тех животных, печень которых обнаруживает ясное дольчатое строение (свинья, медведь, верблюд). У других млекопитающих и у человека в печени так мало соединительной ткани, что она заметна только по ходу сосудов; дольки здесь никогда не бывают ясно очерчены (рис. 396). Тем не менее дольчатое строение имеется и притом такое же, как в печени свиньи и других упомянутых выше животных.
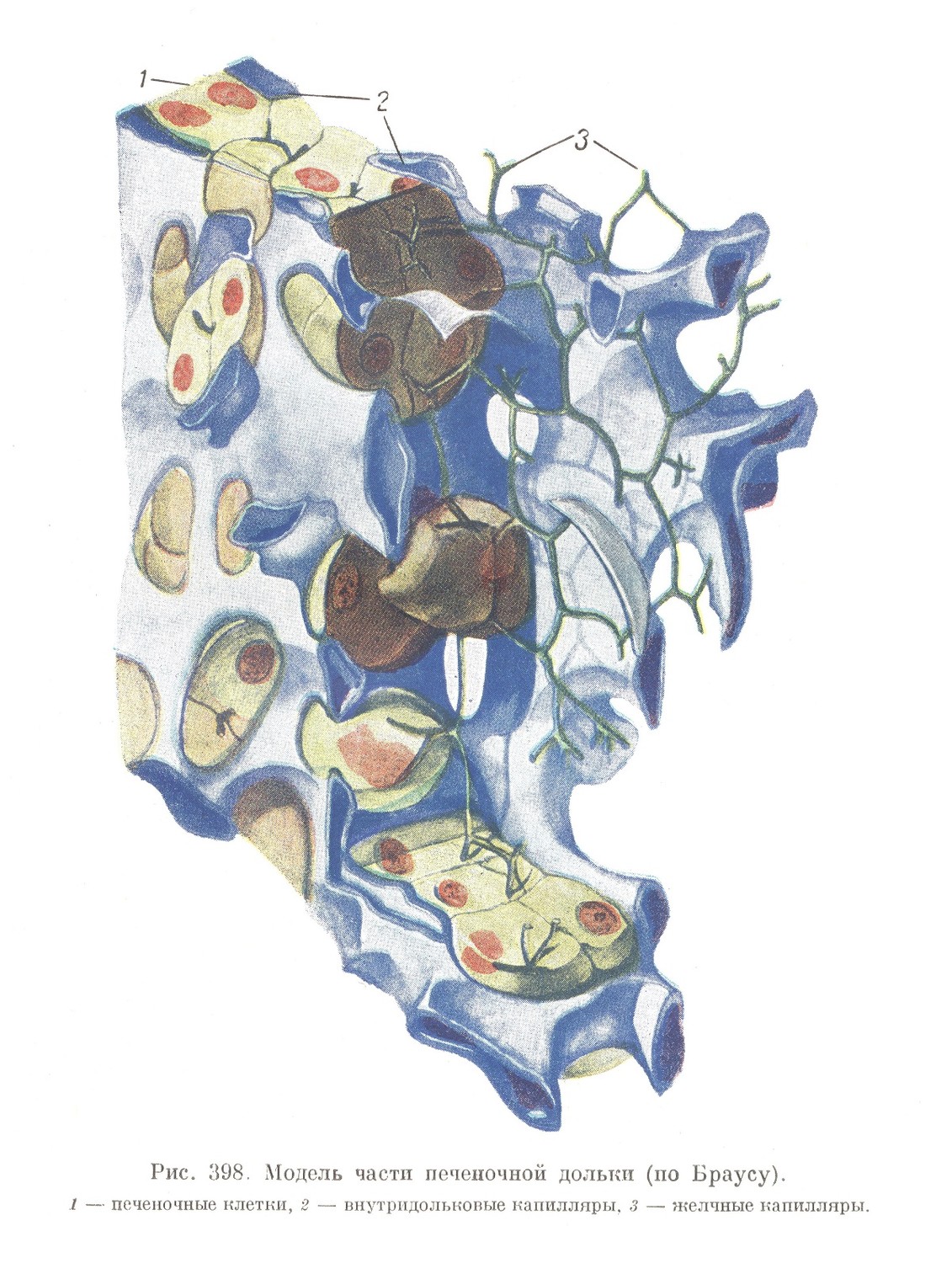
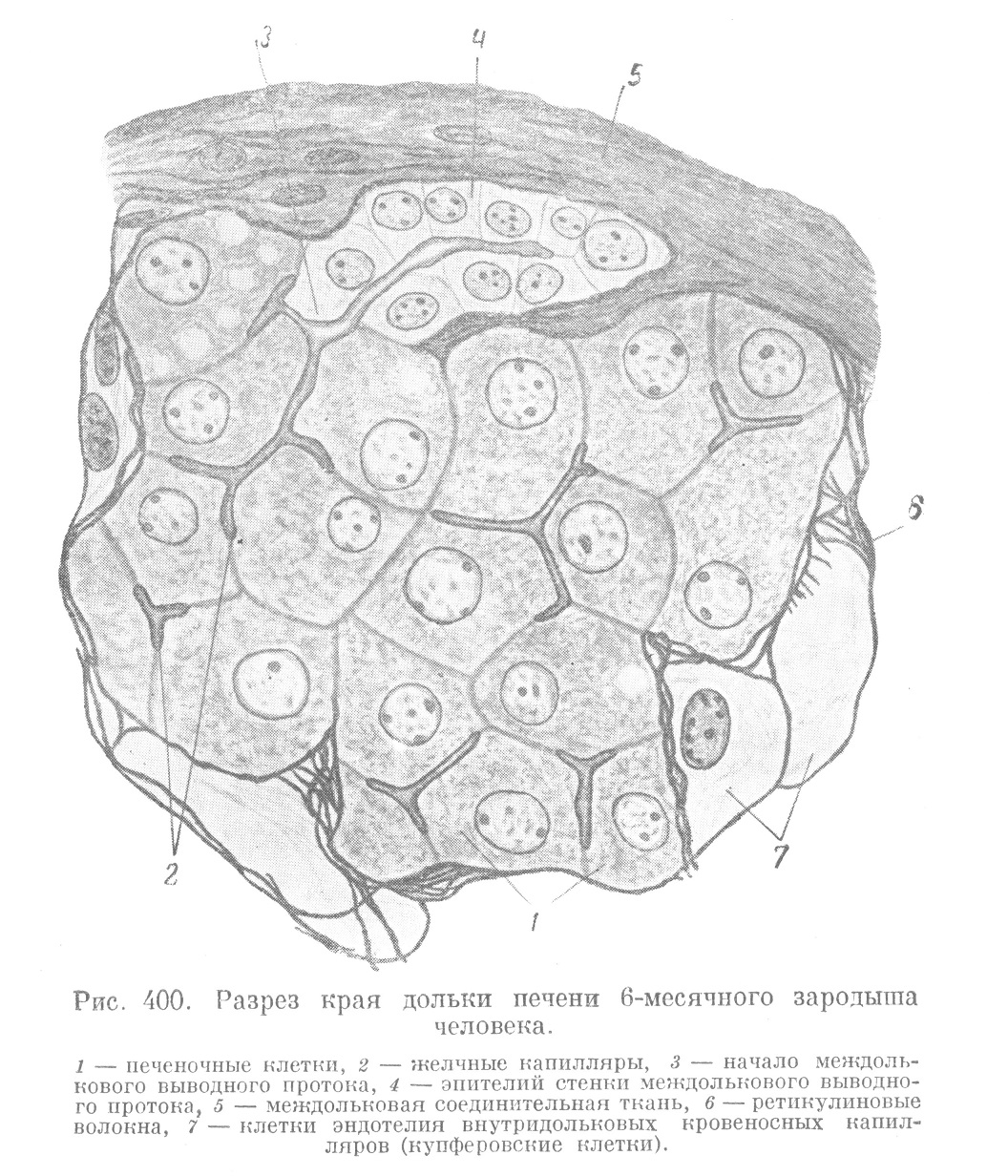
Второе изменение, происходящее при переходе зародышевой печени во взрослое состояние, заключается в том, что эпителиальные тяжи, составляющие ее железистый остов, частично преобразуются в выводные протоки, а частью — в концевые отделы. При этом в выводные протоки превращаются те тяжи, которые оказываются расположенными между образовавшимися дольками. Они в значительной мере утрачивают сетчатый характер и превращаются в каналы, имеющие просвет и эпителиальную стенку. Участки печеночной паренхимы, оказавшейся внутри долек, сохраняют сетчатый характер и преобразуются в своеобразные концевые отделы печени, которые называются печеночными балками (рис. 396). Трубчатый характер и настоящий просвет печеночные балки у млекопитающих и человека имеют только на ничтожном протяжении, а именно в тех частях, которые переходят на периферии долек в междольковые выводные протоки (рис. 400). Вся главная масса печеночных балок настоящих просветов лишена. Вместо них в печеночных балках проходит сеть межклеточных секреторных капилляров (рис. 398 и 399), связанных с упомянутыми периферическими отделами, открывающимися в выводные протоки. У низших позвоночных (например у амфибий) все секреторные отделы имеют трубчатое строение и настоящие, хотя и очень узкие, просветы.

Образовавшиеся дольки имеют форму довольно высоких призм с плоским основанием и выпуклой вершиной. Часто такие простые дольки своими основаниями сливаются между собой (по две и более) в сложные дольки. После всех этих преобразований печень приобретает окончательное строение, изучение которого удобнее всего начать с распределения в ней сосудов.
В ворота печени входят воротная вена и печеночная артерия. Из ворот печени выходят печеночный проток (d. hepaticus) и лимфатические сосуды. В толще печени все разветвления этих сосудов идут вместе в виде общего комплекса по прослойкам соединительной ткани вплоть до печеночных долек. Подойдя к последним, венозные стволики распространяются вдоль их боковых поверхностей в виде так называемых междольковых вен.
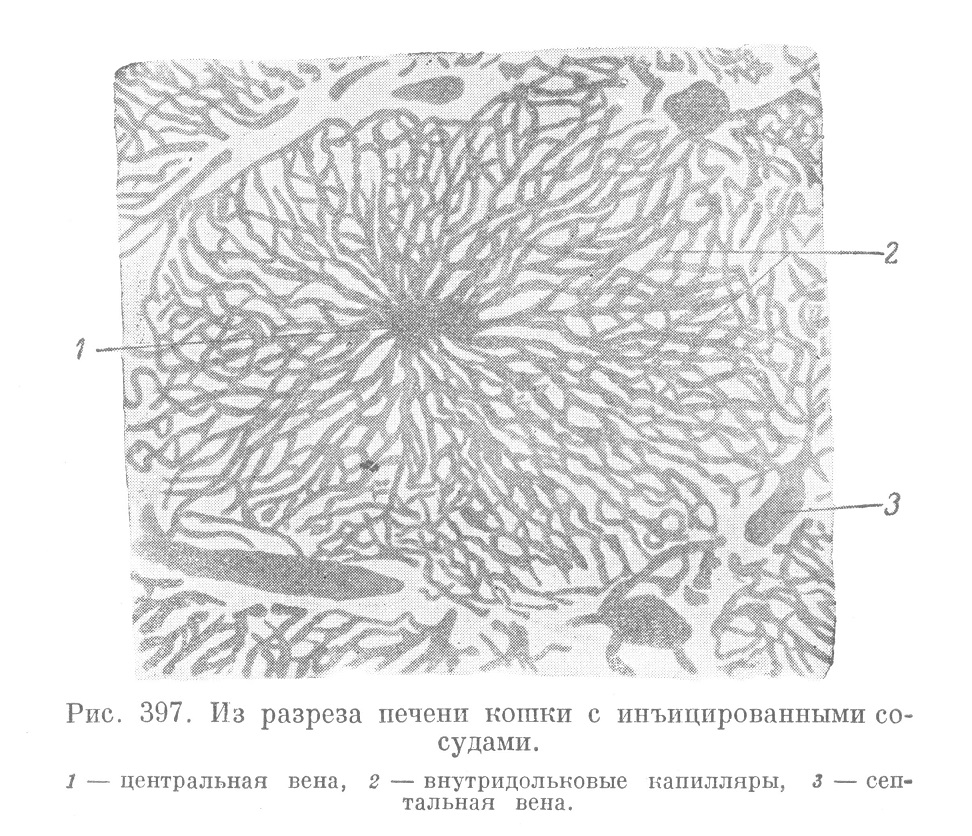
Разветвления последних, называемые септальными венами, окружают каждую дольку со всех сторон.
От септальных вен начинаются широкие капиллярные сосуды (рис. 397 и 398), которые располагаются уже внутри дольки и образуют сеть, выполняющую все промежутки между печеночными балками. Печеночные балки имеют в дольках печени преимущественно радиальное расположение, что связано с основным также преимущественно радиальным направлением сосудов внутридольковой капиллярной сети. В осевой части дольки внутридольковые капилляры сливаются вместе в так называемую центральную (вернее, осевую) вену, (ѵ. centralis) (рис. 397), которая дает начало системе разветвления печеночных вен. Таким образом, сеть внутри дольковых капилляров, омывающих печеночные балки и несущих необходимые для их функционирования вещества, оказывается вставленной между двумя венами.
Из центральной вены кровь переходит, в случае простой дольки, непосредственно в так называемую собирательную вену. В тех случаях, когда центральные вены образуются в частях сложной дольки, они, прежде чем попасть в собирательную вену, сливаются вместе в общий для сложной дольки сосуд, называемый вставочной веной. Эта вена и открывается в собирательную.
Собирательные вены являются разветлениями печеночных вен. Характерным признаком их является то, что они идут по соединительнотканным прослойкам всегда изолированно, не сопровождаемые никакими другими сосудами, что всегда имеет место для разветвленной портальной вены.
Что касается незначительной по размерам печеночной артерии (а. hepatica), то она своими разветвлениями сопровождает разветвления портальной вены и желчных протоков, отдавая к ним мелкие веточки, питающие их стенки. Концевые разветвления печеночной артерии входят и внутрь дольки, но недалеко от ее поверхности впадают в венозную капиллярную сеть.
Несмотря на незначительные размеры, роль печеночной артерии велика. Правда, все вещества, необходимые для питания и работы печеночных клеток, имеются в венозной крови, откуда эти клетки их и черпают, но венозная кровь бедна кислородом, который приносится артериальной кровью. Без этого кислорода печеночные клетки существовать не могут, и перевязки печеночной артерии или тромбоз какой-либо ее веточки вызывают весьма тяжелые нарушения питания, доходящие до омертвения.
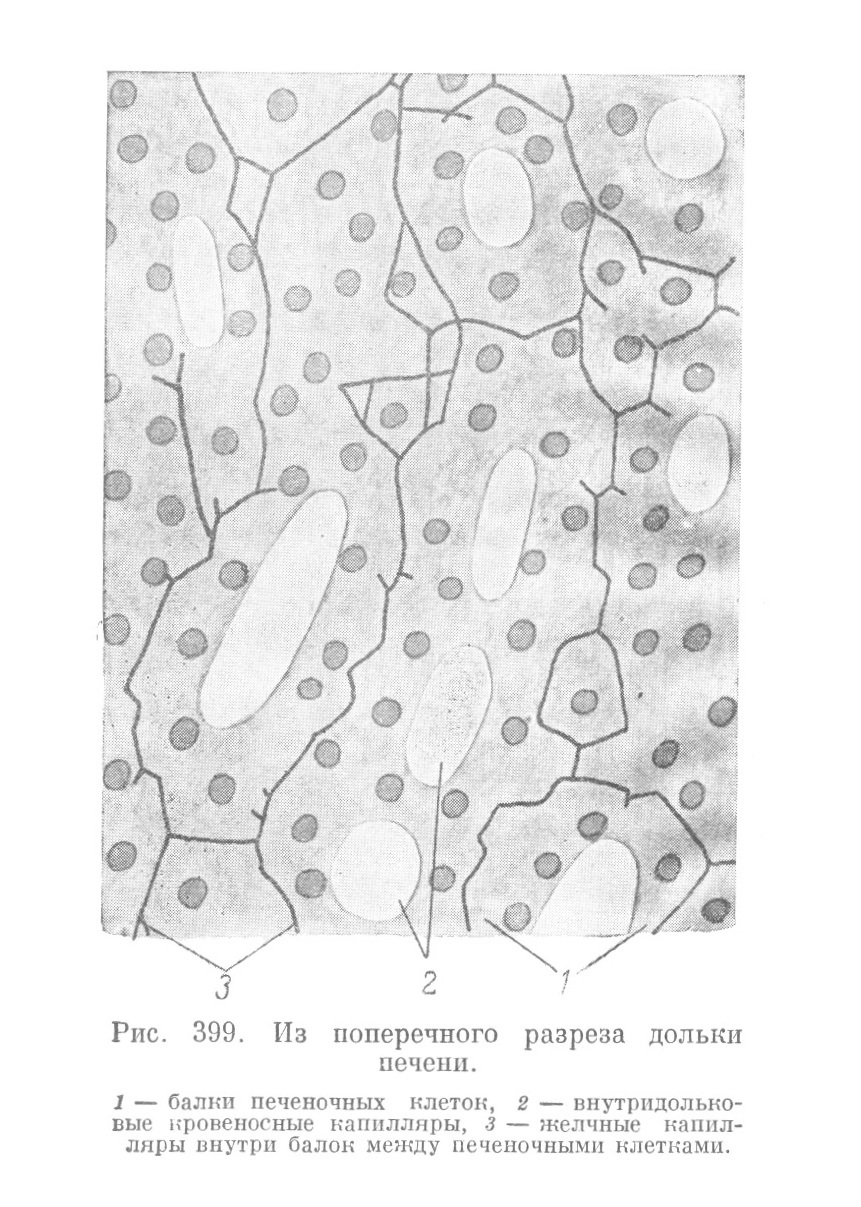
Железистая паренхима печени представлена сетью концевых секреторных отделов, выполняющих все промежутки между капиллярами и называемых печеночными балками. В сети печеночных балок господствует то более (у кролика), то несколько менее (у человека) выраженное радиальное расположение перекладин (рис. 396). Печеночным секреторным отделам у высших позвоночных дают название печеночных балок потому, что, как уже было упомянуто, их структура резко отличается от строения концевых отделов экзокринных желез тем, что балки не имеют настоящего просвета, а секрет в них выделяется в сущности в межклеточные секреторные капилляры. На поперечном разрезе каждая печеночная балка большей частью представляется состоящей всего из 2 клеток (рис. 398). Между ними и проходит секреторный желчный капилляр, просвет которого оказывается образованным двумя желобками на соприкасающихся поверхностях обеих печеночных клеток. Следовательно, в местах соприкосновения любых 2 печеночных клеток всегда находится просвет желчного капилляра. Отсюда ясно, что два просвета могут быть образованы всего тремя клетками, четыре просвета — четырьмя клетками и т. д.
Анастомозирующие тяжи печеночных клеток, образующих печеночные балки, содержат непрерывную сеть также анастомозирующих между собой секреторных желчных капилляров, являющихся межклеточными канальцами, а не настоящими просветами железистых трубок. Настоящие просветы обнаруживаются только в самых периферических частях долек, на месте перехода печеночных балок в междольковые выводные протоки. В эти просветы и открывается сеть внутридольковых желчных капилляров.
Желчные капилляры на обычных препаратах остаются незаметными, и печеночные балки кажутся сплошными клеточными тяжами. Для обнаружения секреторных капилляров необходимо либо наинъицировать их окрашенной массой (рис. 399) через желчный проток, либо импрегнировать серебром. На таких препаратах сеть желчных капилляров выступает с большой отчетливостью, и в ней обнаруживаются некоторые детали, представляющие интерес. Желчные капилляры имеют зигзагообразный ход и от них отходят коротенькие боковые ответвления, слепо заканчивающиеся незначительным вздутием. Эти боковые веточки, но всей вероятности, имеют также межклеточное расположение и вдаются между поверхностями клеток, перпендикулярными к их более длинным осям, вдоль которых проходят капилляры.
Цитоплазма печеночных клеток, отграничивающая просвет желчных капилляров, по-видимому, имеет более плотную консистенцию, потому что при очень осторожной мацерации удается иногда изолировать желчные капилляры в виде небольших обрывков.
Печеночные клетки, образующие печеночные балки, имеют значительные размеры и несколько удлиненную полигональную форму. В результате амитоза эти клетки часто содержат по 2 ядра. Такую двуядерность следует, по-видимому, рассматривать как приспособление для увеличения поверхностных отношений между ядром и цитоплазмой. Необходимость этого вызывается, по- видимому, интенсивной работой, выпадающей на долю печеночной клетки.
Цитоплазма печеночных клеток представляется зернистой и содержит хорошо развитый хондриом. Кроме того, в ней обнаруживаются различные включения: гликоген, жир, пигменты и т. д.
Печеночная клетка в различные фазы своей деятельности имеет различный вид. Наиболее легко обнаруживаются в ней изменения, связанные с углеводным обменом.
Уже через несколько (4—5) часов после приема пищи в печеночных клетках обнаруживаются признаки усиленной деятельности. Клетки несколько увеличиваются в объеме, цитоплазма их становится более зернистой, а ядро — более светлым. Через 10—12 часов тело клетки оказывается наполненным блестящими глыбками гликогена. Затем наступает фаза растворения гликогена, обратное превращение его в глюкозу и выделение последней в кровь. Если не было нового приема пищи, то через 24—48 часов гликоген постепенно исчезает из цитоплазмы, и печеночная клетка принимает первоначальный вид.
Как исключение, в печеночной клетке удается обнаружить и жир. В нормальных условиях жировые включения в печеночных клетках долго не сохраняются и переносятся в другие части организма, где откладываются на более продолжительное время. В печеночных клетках наблюдаются также отложения желчных кислот и пигмента в виде желтых зерен. Другие продукты деятельности печеночных клеток (мочевина) в их цитоплазме под микроскопом не обнаруживаются. В периферической части долек, в местах, граничащих с соединительной тканью, встречаются особые темные (краевые) печеночные клетки, цитоплазма которых интенсивно окрашивается основными красками и не содержит включений.
Таким образом, печеночная клетка обладает свойством двусторонней секреции. Часть веществ, поступающих в нее из крови, она выделяет в желчные капилляры в виде желчи, а все остальные выделяет обратно в кровь. Механизм этой деятельности до последнего времени остается невыясненным. В случае закупорки желчных путей накапливающаяся в дольках желчь попадает, в конце концов, также в кровь и вызывает явления желтухи, но, по-видимому, путем прорыва желчных капилляров, а не путем секреции.
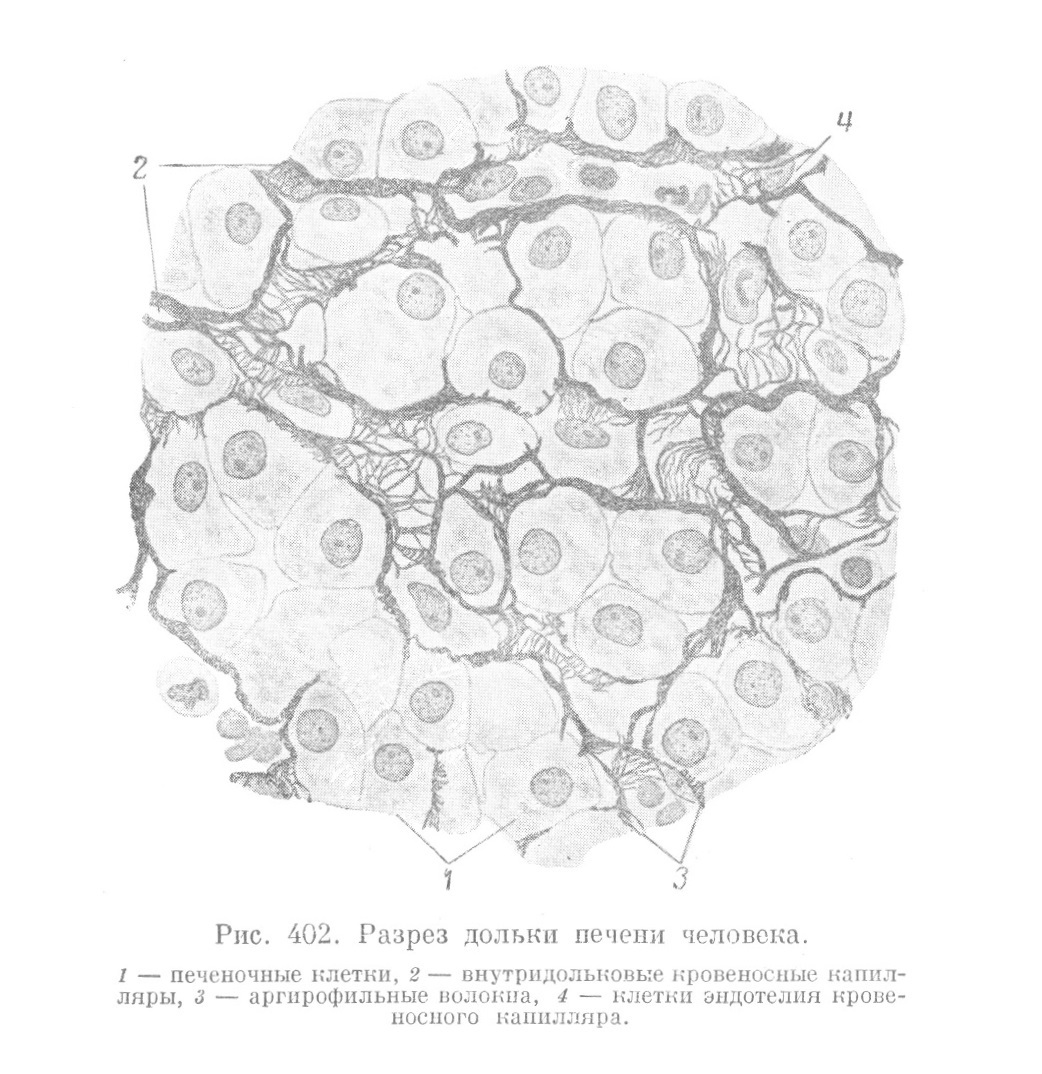
Из сказанного ясно, что отношения печеночных клеток к внутридольковым капиллярам представляют большой интерес и отличаются рядом особенностей. Печеночные балки лишены базальной мембраны, и печеночные клетки прилегают непосредственно к стенке капилляров (рис. 401). Внутри печеночных долек обнаруживаются только решетчатые волокна (рис. 402), образование которых за отсутствием других клеточных элементов приходится отнести за счет эндотелия сосудов.
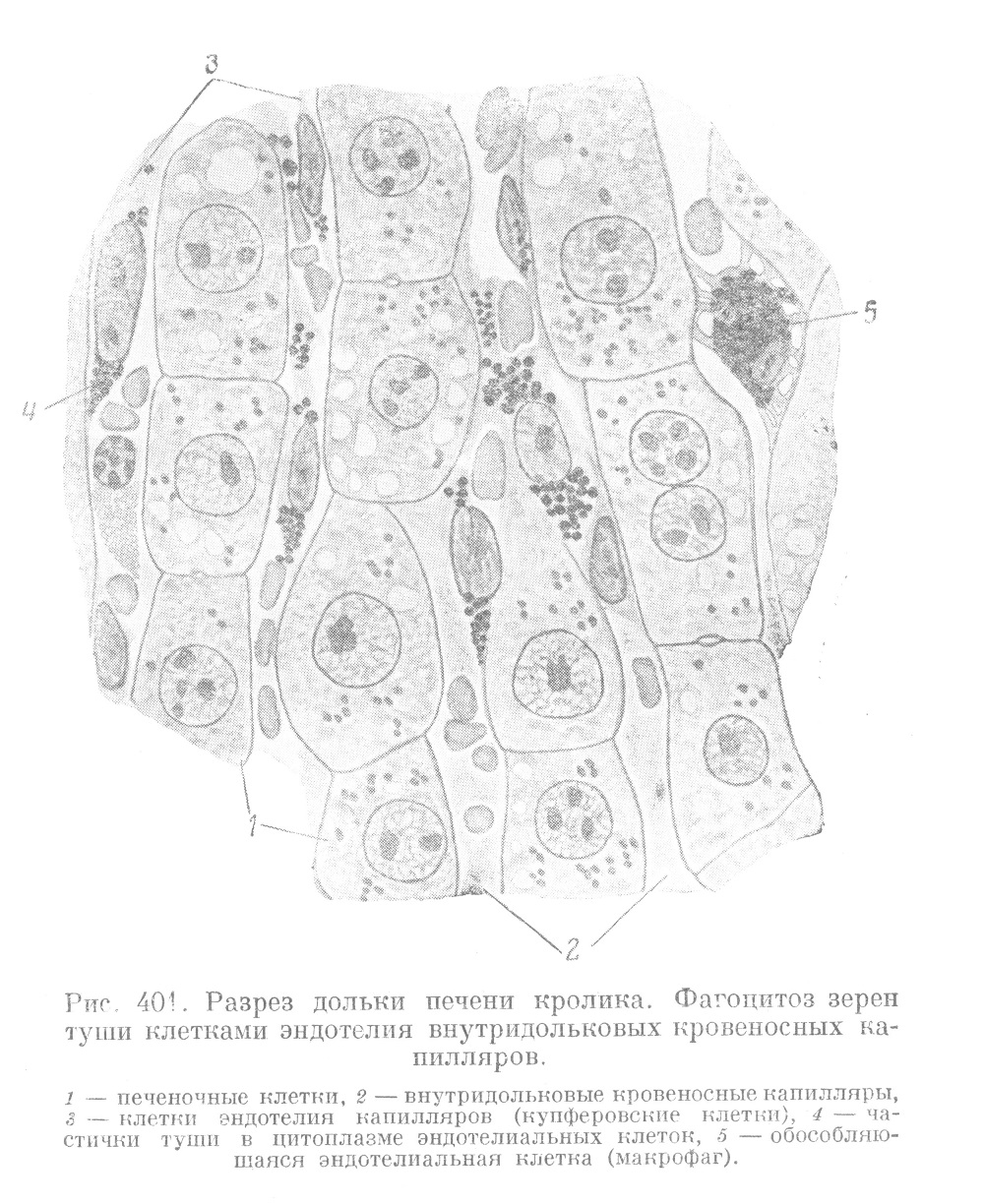
Эндотелий печеночных капилляров имеет своеобразное строение. В печеночных капиллярах обнаружить клеточные границы при помощи метода серебрения не удается. Стенка капилляров представляется состоящей из тонкой бесструктурной пленочки, на которой располагаются синцитиально связанные между собой звездчатые клетки, известные под названием купферовских (рис. 401). Содержащие ядро части этих клеток вдаются в просветы капилляров и омываются протекающей кровью. Купферовские клетки, эти единственные производные мезенхимы, лежащие внутри долек, значительно отличаются от клеток обычного капиллярного эндотелия. Они являются элементами, образующими ретикулиновые (решетчатые) волокна, и обладают всеми теми же свойствами, какие присущи ретикулярным клеткам синусов лимфатических узлов. Купферовские клетки способны к поглощению из крови коллоидных веществ (коллоидное серебро, витальные краски и т. п.) и твердых частиц (эритроциты, бактерии, жир), а также к их перевариванию (фагоцитоз) (рис. 401). Они способны отделяться от стенки и в виде свободных амебоидных элементов примешиваться к элементам крови и т. д. Таким образом, купферовские клетки обладают всеми свойствами элементов ретикуло-эндотелиальной системы, а печень в целом (если принять во внимание ее большой объем) является мощным органом этой системы.
Купферовские клетки в печени несут двоякую функцию. Во-первых, они служат несомненными посредниками между кровью и печеночными клетками: захватывая из крови различные вещества, они передают их печеночным клеткам. Во-вторых, благодаря способности к фагоцитозу и выделению коллоидных веществ они выполняют защитную роль. С этой стороны внутридольковые печеночные капилляры очень похожи на синусы лимфатических узлов, селезенки или костного мозга.
Выводные протоки печени начинаются мелкими разветвлениями междольковых желчных протоков, сообщающимися указанным выше способом с внутридольковыми желчными капиллярами (рис. 400). Они имеют простое строение. Стенка их состоит из однослойного кубического эпителия, расположенного на бесструктурной мембране. Клетки этого эпителия имеют кутикулярную каемку. Более крупные желчные протоки проходят вместе с разветвлениями портальной вены и почечной артерии. Эти протоки имеют уже собственную соединительнотканную стенку и выстланы призматическим эпителием, который по мере увеличения калибра протоков становится все более и более высоким. Главные выводные протоки d. hepaticus, d. cysticus имеют хорошо выраженный соединительнотканный собственный слой с большим количеством эластических волокон. За собственным слоем следует незначительный слой кольцевых мышц, к которым на месте впадения в кишку присоединяются и продольные мышечные слои; последние, сливаясь с мышечной стенкой кишки, образуют нечто вроде сфинктера. Большие выводные протоки выстланы особенно высоким призматическим эпителием, который, повидимому, имеет секреторный характер. В цитоплазме его клеток обнаруживаются базофильные гранулы, постепенно превращающиеся в особый муцин, который выделяется в просвет протока и примешивается к желчи. В больших желчных протоках наблюдаются так называемые железы желчных путей.
В печени обнаруживаются еще и слепо заканчивающиеся желчные протоки, расположенные вне печеночной паренхимы. Они встречаются, главным образом, на левом краю печени, в воротах и в области нижней полой вены. D. aberrantes не что иное, как редуцированные остатки железистой паренхимы эмбриональной печени.
Стенка желчного пузыря состоит: 1) из собранной в высокие складки слизистой оболочки, 2) мышечной оболочки и 3) наружной фиброзной оболочки, которая со стороны брюшной полости покрыта серозным покровом.
Слизистая оболочка выстлана высоким призматическим эпителием, содержащим в одних клетках базофильные, в других — оксифильные гранулы. В эпителии содержатся также липоиды. Слизистая оболочка его в функциональном отношении отличается тем, что через нее происходит всасывание воды из находящейся в полости желчи. Таким образом, желчь в пузыре имеет более густую консистенцию, чем желчь, изливающаяся из печени.
Мышечная оболочка желчного пузыря состоит, главным образом, из циркулярно расположенных мышечных пучков, к которым (в меньшем числе) присоединяются и продольные и косые мышечные пучки.
Нервы печени относятся, главным образом, к сосудистым вегетативным нервам; кроме того, в печени, по-видимому, имеются и секреторные нервы, заканчивающиеся на печеночных клетках. Из двенадцатиперстной кишки в выводные протоки печени заходят участки ауэрбахова сплетения.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 