Эмбриональное развитие высших позвоночных отличается от эмбриогенеза низших позвоночных животных прежде всего тем, что оно происходит на суше. В одних случаях оно протекает свободно, и тогда зародыш бывает окружен специальными оболочками (скорлупа, скорлуповая оболочка яиц рептилий, птиц и яйцекладущих млекопитающих). В других случаях это развитие протекает в половых путях материнского организма. Указанные внешние условия значительно изменяют процесс развития высших позвоночных, которое у всех представителей этой группы, а равно и у человека является меробластическим.
Во всех случаях у зародыша очень рано развиваются временные провизорные органы различного строения и функции. Среди них следует отметить один постоянно встречающийся провизорный орган, создающий водную среду для развивающегося зародыша — амнион. Этот орган образуется у всех высших позвоночных животных и человека. На основании этого признака их принято объединять в одну группу — амниот, в противоположность низшим позвоночным животным, лишенным амниона и составляющим группу — анамниа.
Особенности строения зародыша высших позвоночных, связанные с развитием провизорных органов, относятся к ценогенетическим признакам, которые отражают приспособительную реакцию зародыша к условиям внешней среды. Последние весьма изменчивы, в связи с чем и провизорные органы имеют различное строение.
Развитие птиц
Примером подобного развития может служить эмбриогенез цыпленка. Яйцевая клетка курицы является телолецитальной. Она содержит значительное количество желтка и вследствие этого приобретает гигантские размеры. Оплодотворение у птиц — внутреннее, и совершается оно в половых путях материнского организма. Строение зиготы остается еще неизученным. Дробление — частичное или меробластическое. Оно происходит во время движения зародыша по яйцеводу в сторону клоаки. Первая меридиональная борозда разделяет зиготу в области анимального полюса на два бластомера. Последние остаются соединенными своими основаниями с неразделившейся частью зиготы.
Вторая меридиональная борозда, как и первая, проходит на анимальном полюсе и разделяет каждый из бластомеров на два новых. Образуется зародыш из четырех бластомеров. Третья борозда — широтная. В дальнейшем наблюдается чередование меридиональных, широтных и тангенциальных борозд. В процессе дроблении количество бластомеров в составе зародыша быстро увеличивается, а их размеры с каждым делением уменьшаются.
Движение развивающегося зародыша сопровождается реакцией со стороны слизистой оболочки яйцевода. Она проявляется в том, что железистый аппарат слизистой оболочки выделяет большое количество белкового секрета, который связывается с поверхностью зародыша, обволакивая его толстым слоем. Позднее снаружи от слоя белкового секрета возникает тонкая плотная пленка (скорлуповая оболочка), состоящая из двух соединенных вместе листков.
В нижних отделах половых путей, снаружи от скорлуповой оболочки, откладывается твердая известковая оболочка — скорлупа. Подобно белку, скорлуповая оболочка и скорлупа являются продуктами жизнедеятельности половых путей. В таком виде зародыш птиц, будучи окруженным указанными выше оболочками, откладывается во внешнюю среду и в целом называется яйцом.
Так как температура тела птицы значительно выше температуры внешней среды, то отложенное яйцо охлаждается. Вместе с этим приостанавливается и развитие зародыша. Развитие прерывается в период дробления, которое еще не завершилось. В таком состоянии зародыш пребывает до тех пор, пока внешние условия не станут благоприятными для его дальнейшего развития, которое может быть вызвано или насиживанием яиц птицей, или помещением яиц в инкубатор.
Через два часа пребывания зародыша в инкубаторе период дробления заканчивается. На анимальном полюсе расчлененной части зиготы образуется дискобластула. Она состоит из наружного и внутреннего слоев» которые переходят друг в друга по краю дискобластулы.
Полость дискобластулы (бластоцель) представляет сложную щель между наружным и внутренним слоями (рис. 56, Б). Внутренний слой состоит из крупных элементов, расположенных в один ряд и содержащих большое количество желточных зерен. В одних элементах ядра отчетливо выступают, в других — их трудно заметить. В последнем случае они носят название желточных шаров. По О. Б. Лепешинской, желточные шары представляют элементы живого вещества, не имеющие клеточного строения.
Внутренний слой дискобластулы, непосредственно связанный с общей нерасчлененной желточной массой, выполняет очень важную функцию, резорбируя желток (а в дальнейшем и белок), как питательный материал. На это еще в свое время указывал Ф. Энгельс: «...обыкновенный белок играет наиболее безжизненную, наиболее пассивную роль: наряду с желтком, белок служит исключительно питательным веществом для развивающегося зародыша» (Ф. Энгельс. Анти-Дюринг, стр. 77, 1950).
Наружный слой дискобластулы образован более мелкими клетками, которые по сравнению с элементами внутреннего слоя содержат желтка меньше.
Если рассматривать дискобластулу в естественном ее положении, с наружной поверхности, то она имеет округлую форму. В ней различают небольшую центральную часть, представленную зародышевым щитком (рис. 56, А). Вокруг него широкой полосой, приподнятой слегка над желтком и поэтому кажущейся более светлой, расположено светлое поле. Последнее окружено широкой циркулярной полосой, называемой темным полем. Темное поле переходит в край обрастания, который вместе с тем является и краем дискобластулы.
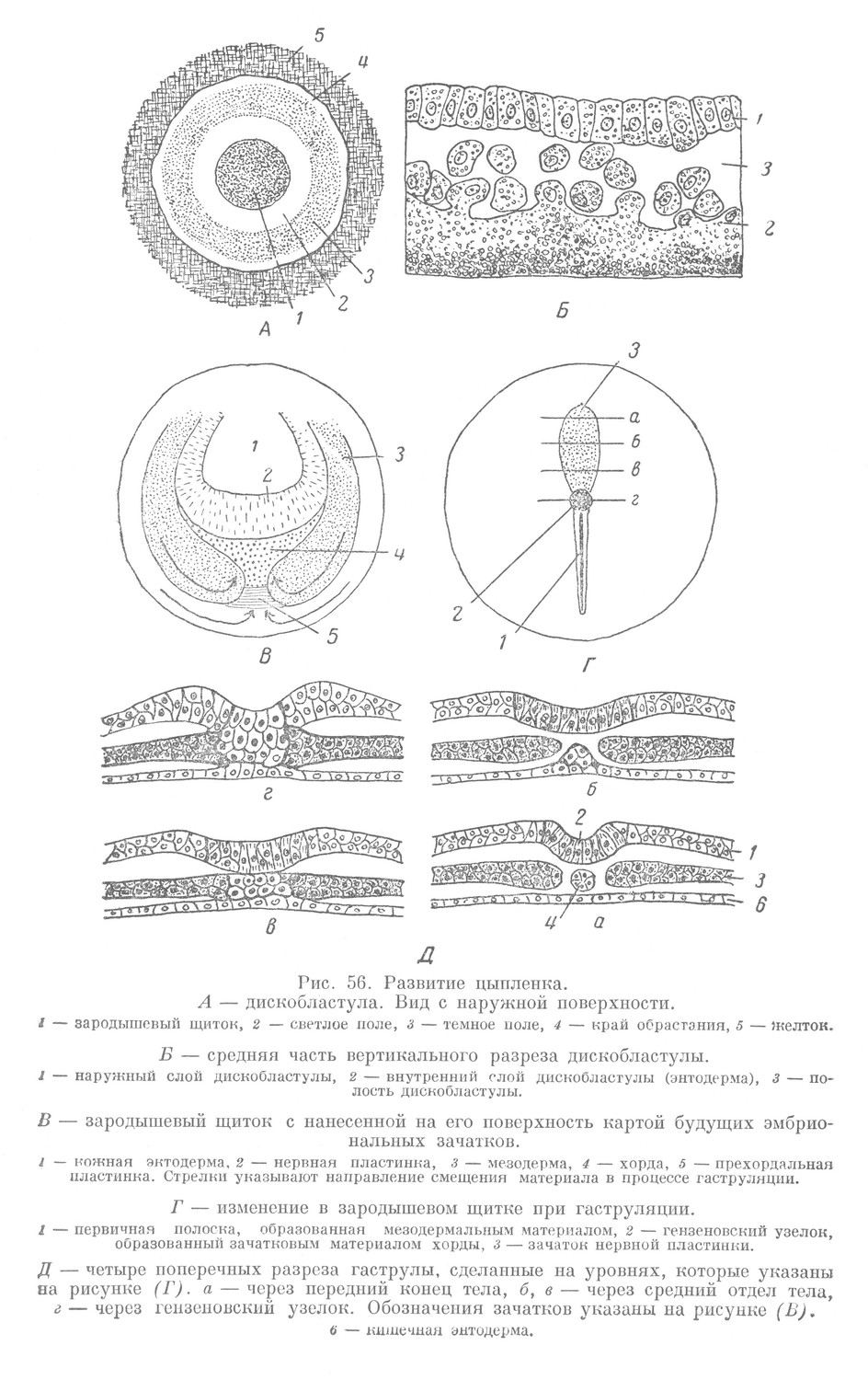
Зародышевый щиток представляет ту часть эмбриона, которая дает начало зачаткам окончательных органов его тела, т. е. является зародышевым материалом. Вся остальная значительная по размерам часть дискобластулы, образованная светлым и темным полями и краем обрастания, составляет внезародышевый материал, служащий источником образования всех провизорных органов.
Сравнивая между собой дискобластулы птиц и рыб, нетрудно отметить различия (рис. 51, а, 56, А). Зародышевый туштедиал в дискобластуле рыб находится в краевом положении, тогда как в дискобластуле птиц — в срединном. Количество внезародышевого материала в дискобластуле птиц преобладает над количеством зародышевого, у рыб же основную часть дискобластулы составляет зародышевый материал.
В составе зародышевого щитка находится материал будущих зачатков, которые четко выявляются в период гаструляции. Расположение предполагаемых зачатков у птиц такое же, как и в бластуле ланцетника, миноги, в зародышевой части дискобластулы рыб и в бластуле амфибий (рис. 56, В).
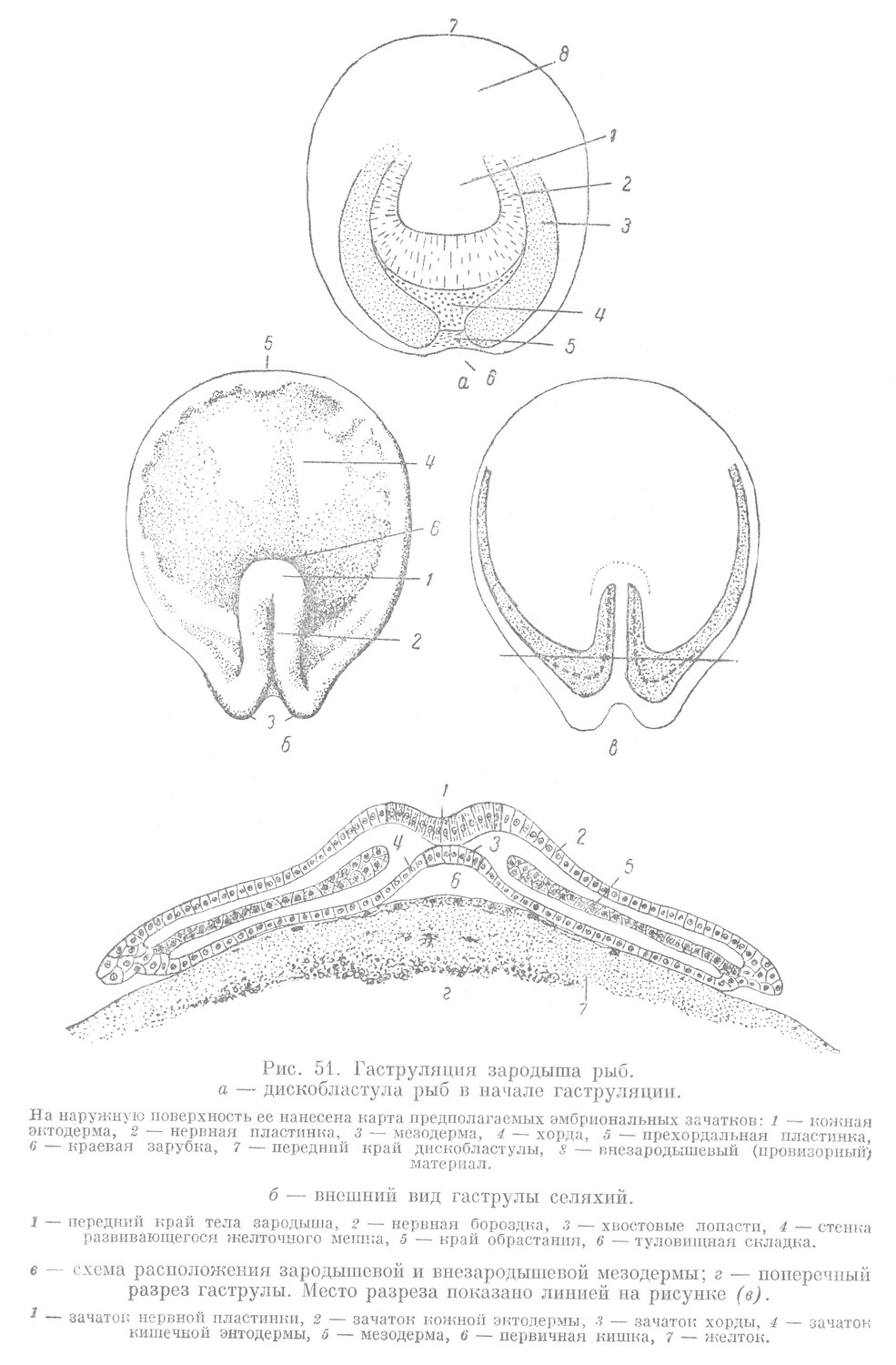
Гаструляция у птиц, так же как и у низших позвоночных, характеризуется двумя основными чертами. С одной стороны, здесь, вследствие неравномерного роста, наблюдается определенный сдвиг отдельных участков в составе зародышевого щитка, с другой стороны — их диференцировка. Перемещение материала приводит к образованию в задней половине зародышевого щитка валика, идущего вдоль зародыша, который называют первичной полоской (рис. 56, Г). У переднего конца первичной полоски образуется утолщение, называемое гензеновским узелком. В ходе дальнейшей гаструляции от гензеновского узелка в сторону бластоцеля и по направлению к переднему концу тела зародыша под наружным слоем вырастает в виде тяжа головной, или хордальный, отросток. Он является зачатком хорды.
От первичной полоски по бокам хорды, также в сторону переднего конца тела зародыша, вырастают два крыла, образующие зачаток мезодермы. В наружном слое в виде полоски над хордальным отростком располагается зачаток нервной пластинки. Наиболее глубоко располагается зачаток кишечной энтодермы. Он непосредственно прилегает к поверхности желтка и находится во внутреннем слое дискобластулы в области расположения зародышевого щитка вместе с прехордальной пластинкой, лежащей у переднего конца зародыша.
Из сказанного следует, что к концу гаструляции в теле зародыша выявляются все зачатки, расположенные в соответствии с принципом осевого строения позвоночных (рис. 56, Д). Этот признак отмечался также и у низших позвоночных животйых и относится к числу палингенетических. В деталях взаимоотношения эмбриональных зачатков имеют, однако, свои особенности. Прежде всего основной зачаток, входящий в состав стенки первичной кишки низших позвоночных — кишечная энтодерма, у птиц очень рано обособляется от материала будущих основных зачатков (хорды и мезодермы). Уже в конце дробления он оказывается отделенным от них и находится в составе внутреннего слоя дискобластулы на уровне расположения зародышевого щитка. Следовательно, у птиц обособление материала будущих зачатков кишечной энтодермы, хорды и мезодермы происходит раньше, чем у амфибий, а именно в период дробления при образовании дискобластулы (рис. 56, В).
Сравнивая строение гаструлы всех ранее рассмотренных низших позвоночных животных с гаструлой птиц, нетрудно установить, что строение гаструлы птиц есть дальнейший этап усложнения эмбриогенеза позвоночных животных. Хорда и мезодерма у птиц полностью утрачивают связь со стенкой первичной кишки, а выделение кишечной энтодермы у них происходит значительно раньше, чем у амфибий.
Развитие внезародышевого материала у птиц идет одновременно с развитием зародышевого, однако скорость его диференцировки значительно опережает развитие тела зародыша. На это указывалось раньше, когда излагался эмбриогенез рыб, у которых появление тканей при развитии желточного мешка происходит значительно быстрее, чем развитие тканей в теле зародыша. У птиц рост и диференцировка провизорного материала происходит более быстрыми темпами по сравнению с рыбами. Уже в конце дробления провизорный материал доминирует над материалом, являющимся источником, из которого развиваются дефинитивные органы. Разрастание материала провизорных органов идет равномерно во все стороны по поверхности нерасчлененной части зиготы, при этом раньше всего выявляется развитие желточного мешка — органа трофической функции. Разрастающаяся провизорная часть зародыша вначале состоит из наружного слоя, представленного внезародышевой эктодермой, внутреннего слоя, образованного желточной энтодермой, и среднего слоя, представленного двумя листками мезодермы: париетальным и висцеральным.
После завершения гаструляции и в результате роста туловищной складки начинается обособление тела зародыша от провизорного материала. Туловищная складка у птиц имеет то же направление, что и у рыб. Начинается она в области головного конца тела зародыша и постепенно распространяется по всей окружности в каудальном направлении, отделяя тело зародыша от желточного мешка.
Одновременно с появлением туловищной складки и несколько кнаружи от нее появляется амниотическая складка (рис. 57, а). Она образована внезародышевой эктодермой и париетальным листком внезародышевой мезодермы. Направление ее роста противоположно направлению туловищной складки, в результате чего амниотическая складка нарастает на тело зародыша. Растущие навстречу друг другу края амниотической складки сближаются и срастаются, окружая тело зародыша. Это приводит к образованию одновременно двух провизорных органов —- амниона и серозной оболочки (рис. 57).
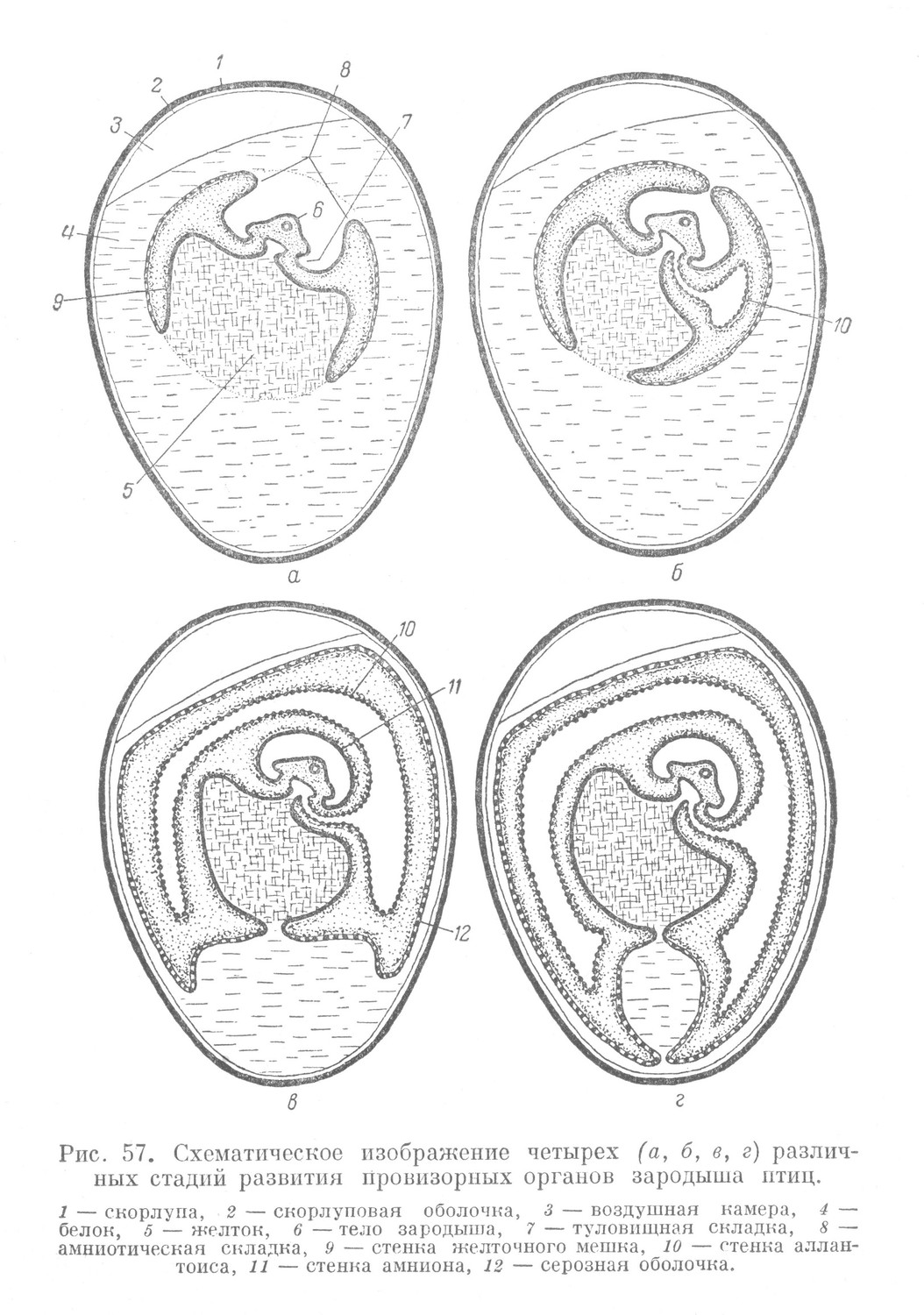
Амниотическая, или водная, оболочка образуется в результате срастания стенок внутренних склонов амниотической складки и состоит из внезародышевых эктодермы и мезодермы. Она ограничивает полость, заполненную жидкостью, в которой пребывает тело зародыша. Амниотическая жидкость является продуктом жизнедеятельности амниотической оболочки.
После срастания краев амниотической складки из стенок наружных склонов образуется серозная оболочка — провизорный орган дыхания зародыша. Ее образуют внезародышевая эктодерма и париетальный листок внезародышевой мезодермы. Этот орган в соответствии с выполняемой им функцией располагается наиболее поверхностно по сравнению с другими провизорными органами, прилегая непосредственно к скорлуповой оболочке и скорлупе, т. е. в наибольшей близости, к окружающему яйцо воздуху.
Одновременно с развитием амниона и серозной оболочки развивается четвертый провизорный орган — аллантоис, имеющий вид мешка. Он возникает частично из кишечной энтодермы путем выпячивания в виде кармана вентральной стенки задней кишки и, покидая тело зародыша, увлекает за собой висцеральный листок внезародышевой мезодермы (рис. 57, б). Аллантоис сильно разрастается и располагается по всей окружности зародыша. Он заполняет щель между желточным мешком и амнионом, с одной стороны, и серозной оболочкой, с другой. Стенка его состоит из кишечной энтодермы, ставшей внезародышевой, и висцерального листка внезародышевой мезодермы.
Указанные выше провизорные органы в ходе развития быстро приобретают тканевую организацию. Часть материала стенки желточного мешка еще в процессе обрастания начинает диференцироваться в ткани. Из желточной энтодермы образуется желточный эпителий — ткань пограничной функции (рис. 58). Диференцировка материала висцерального листка внезародышевой мезодермы одновременно приводит к образованию мезенхимы. В последней развивается сеть кровеносных капилляров, непосредственно прилегающих к базальной поверхности желточного эпителия.
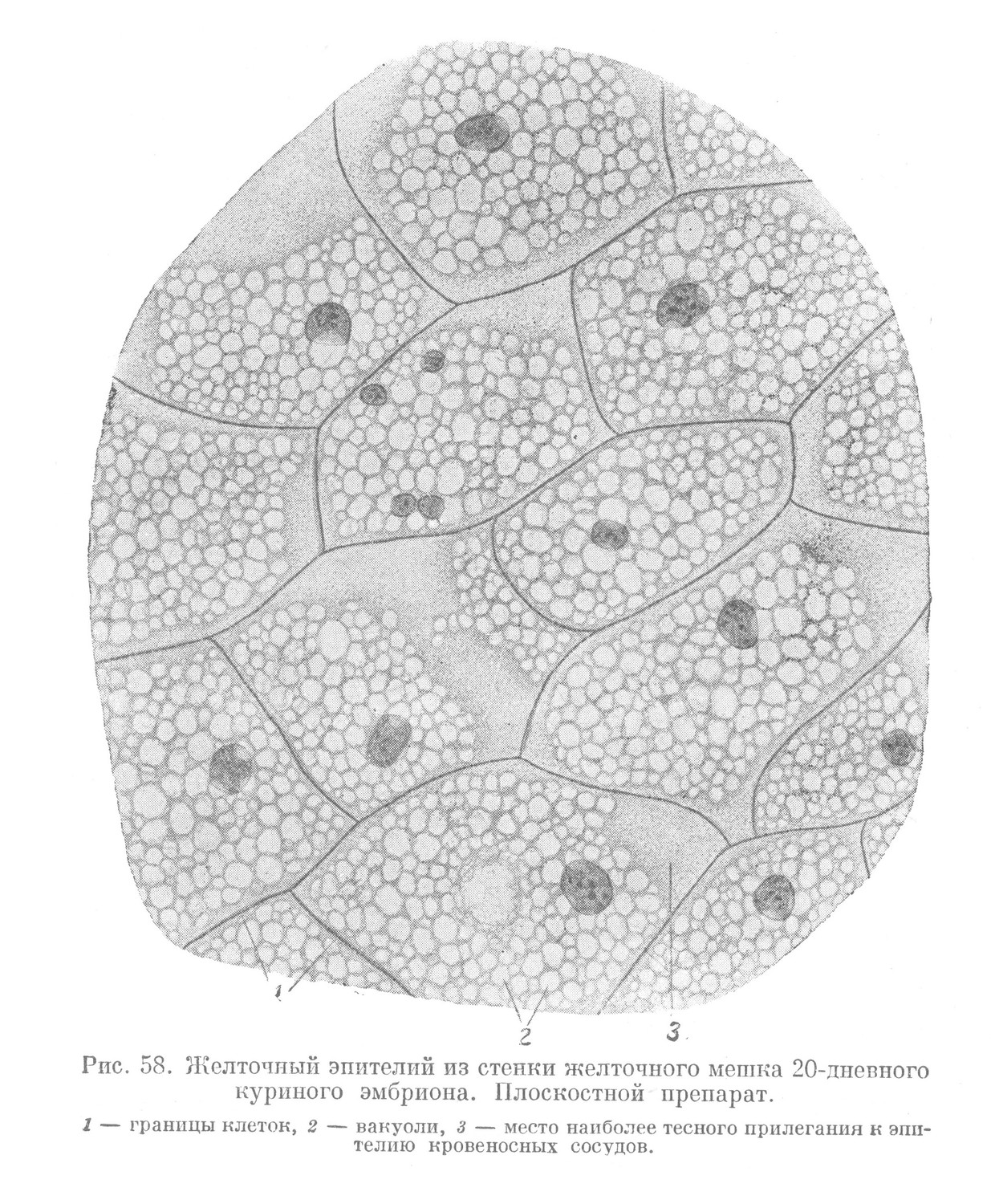
Кровеносные капилляры представляют конечное разветвление желточных сосудов, главные стволы которых возникают в мезенхиме желточного стебелька, направляясь к телу зародыша. Одновременно с сосудами в мезенхиме образуются первые клетки крови. Мезенхиму желточного мешка следует рассматривать как начальный провизорный орган кроветворения.
Зачатковый материал оболочки амниона, вскоре после ее образования, превращается в различные ткани. Из внезародышевой эктодермы возникает амниотический эпителий. Он представляет тонкий пласт, состоящий из плоских клеток, имеющих с поверхности вид полигональных пластинок значительного размера. В эпителии наблюдаются митозы и амитозы. Встречаются также и многоядерные клетки. Амниотический эпителиіі в области канатика переходит в кожную эктодерму тела зародыша, причем граница между ними очень резкая.
Париетальный листок внезародышевой мезодермы одновременно с эктодермой также развивается в ткань — мезенхиму. Развитие последней сопровождается образованием в стенке амниона различного калибра кровеносных сосудов, расположенных в молодой соединительной ткани, возникшей из мезенхимы. Кроме того, из мезенхимы развивается значительный слой гладкой мышечной ткани, образующий мощный сократимый аппарат стенки амниона.
Функцией амниотической оболочки является прежде всего выработка амниотической жидкости, которая служит средой для тела развивающегося зародыша. Химический состав амниотической жидкости более или менее постоянен и зависит от жизнедеятельности амниотической оболочки. Хорошо развитый мышечный слой ее поддерживает в амниотической полости определенное давление. Таким образом, амнион создает необходимую водную среду для развития тела зародыша.
Возникновение амниона в эволюции эмбриогенеза является, несомненно, актом приспособительного характера, который свойственен эмбриогенезу не только высших позвоночных, но и отдельным группам беспозвоночных (насекомые). Это — адаптация зародыша к новым внешним условиям и прежде всего к развитию на суше.
Возникшая одновременно с амнионом серозная оболочка до недавнего времени рассматривалась как лежащая наиболее поверхностно и выполняющая лишь защитную функцию. Однако, несмотря на простоту анатомического строения, в действительности она имеет сложную гистологическую структуру и представляет жизненно важный временный орган зародыша, осуществляющий дыхательную функцию. Этот провизорный орган дыхания, так же как и другие временные органы зародыша, очень рано приобретает тканевое строение. Источником развития образующих его тканей являются два зачатка: внезародышевая эктодерма и париетальный листок внезародышевой мезодермы. Первая дает респираторный или дыхательный эпителий, второй — мезенхиму, развивающуюся в соединительную ткань и кровеносные сосуды. В морфологическом отношении респираторный эпителий весьма сходен с одноименным эпителием дефинитивных органов дыхания позвоночных (эпителий жаберных лепестков и альвеол легких). В подлежащей соединительной ткани серозной оболочки находится узкопетлистая густая сеть кровеносных капилляров (рис. 59).
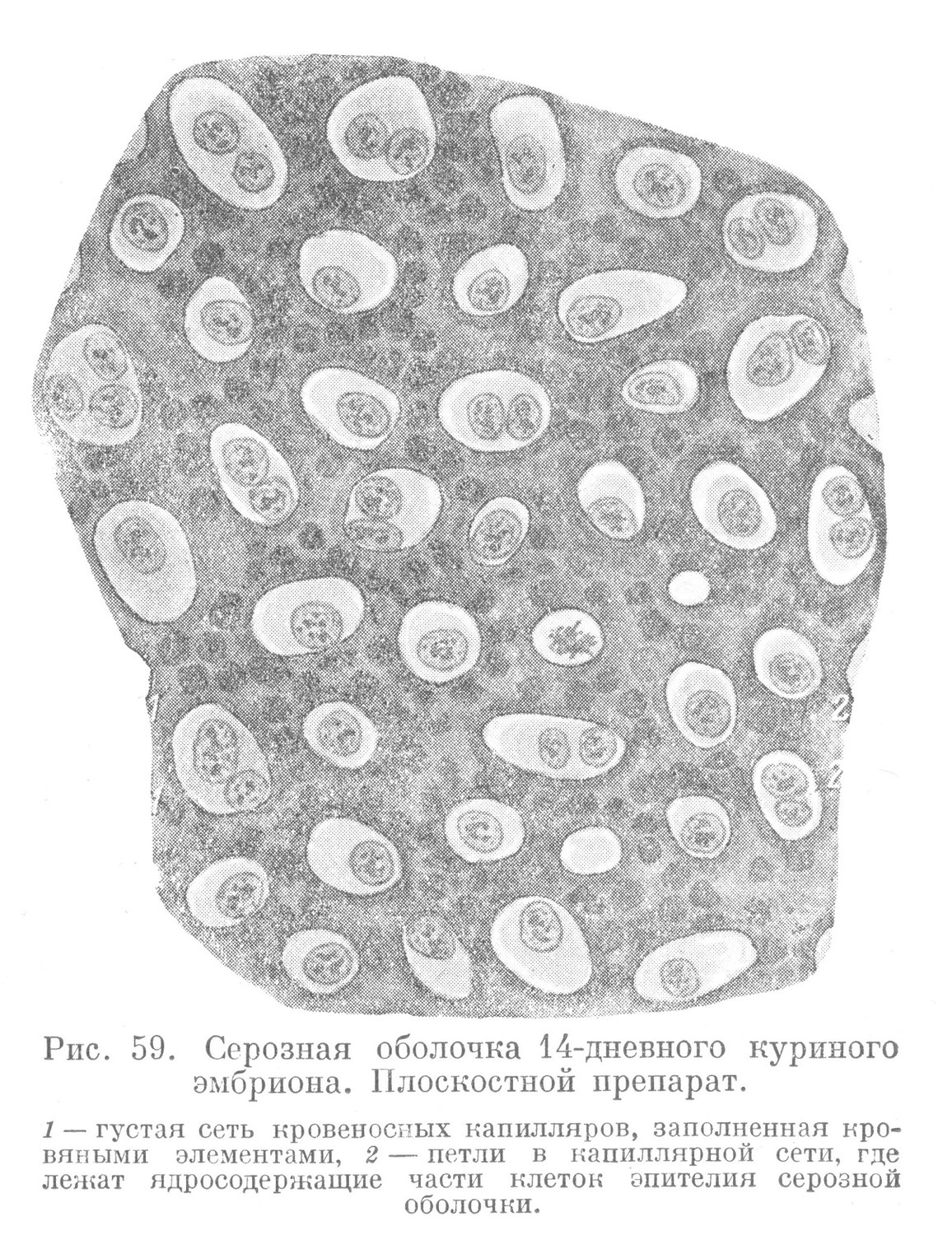
Самой характерной чертой строения дышащей части всех дефинитивных органов дыхания является наиболее тесное и весьма своеобразное взаимоотношение респираторного эпителия с подлежащей сетью кровеносных капилляров. Эта черта и определяет строение серозной оболочки. Поэтому имеются все основания рассматривать ее как провизорный орган дыхания зародыша. Кислород воздуха, проникший через поры скорлупы и скорлуповую оболочку, попадает через респираторный эпителий в кровеносные капилляры и связывается здесь с гемоглобином эритроцитов. Вместе с этим в обратном порядке происходит выделение накопившейся в крови углекислоты. Процесс газового обмена обусловливается различным парциальным давлением названных газов в крови зародыша и воздухе, проникающем через скорлупу. Принимая во внимание большую поверхность серозной оболочки, следует предполагать, что газовый обмен в этих условиях протекает весьма интенсивно.
Материал стенки развивающегося аллантоиса, подобно другим провизорным органам, также быстро превращается в ткани. Внезародышевая энтодерма дает однослойный плоский эпителий, выстилающий полость аллантоиса. (считают, что задняя кишка зародыша, из вентральной стенки которой развивается аллантоис, имеет не энто-, а эктодермальную природу. Вследствие этого развивающиеся из нее эпителиальные ткани (эпителий мочевого пузыря и каудальная часть задней кишки) имеют строение, свойственное энителиям эктодермального типа.)
Висцеральный листок внезародышевой мезодермы превращается в мезенхиму, которая частично сливается с мезенхимой серозной оболочки и участвует в образовании сосудов. В мезенхиме аллантоиса развиваются главные стволы пупочных сосудов, идущих от тела зародыша к сосудистой сети серозной оболочки. Вместе с тем аллантоис несет и функцию выделения, так как в его полости накапливаются продукты азотистого обмена.
Рассмотренные выше внезародышевые образования, которые принято называть зародышевыми оболочками, представляют собой временные органы зародыша. Вместе с последним они составляют целостный развивающийся организм, в котором на первом этапе интегрирующую роль играет кровеносная система. Позднее эта роль в значительной степени переходит к нервной системе, хотя все провизорные органы лишены ее элементов.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 