Развитие речной миноги
Яйцевая клетка миноги содержит значительное количество желтка, расположенного преимущественно у вегетативного полюса. Яйцевые клетки с таким неравномерным распределением желтка, сосредоточенного на вегетативном полюсе, называются телолецитальными. Оплодотворение происходит свободно, в водной среде. Значительное количество желтка в зиготе маскирует происходящие в ней изменения. Вследствие этого они остаются до настоящего времени недостаточно изученными.
Дробление зиготы — полное. Оно начинается с появления первой меридиональной борозды, разделяющей зиготу на два связанные между собой бластомера (рис. 48, а). Борозда возникает на анимальном полюсе и ввиду значительного количества желтка медленно распространяется к вегетативному полюсу. Далее перпендикулярно к плоскости предыдущего деления появляется вторая меридиональная борозда и, подобно первой, делит каждый бластомер на два новых. Возникает зародыш, состоящий из 4 бластомеров (рис. 48, б). Третья борозда — экваториальная — проходит не строго по экватору, а слегка сдвинута в сторону анимального полюса. Образуется стадия 8 бластомеров. Далее следует чередование меридиональных и широтных борозд.
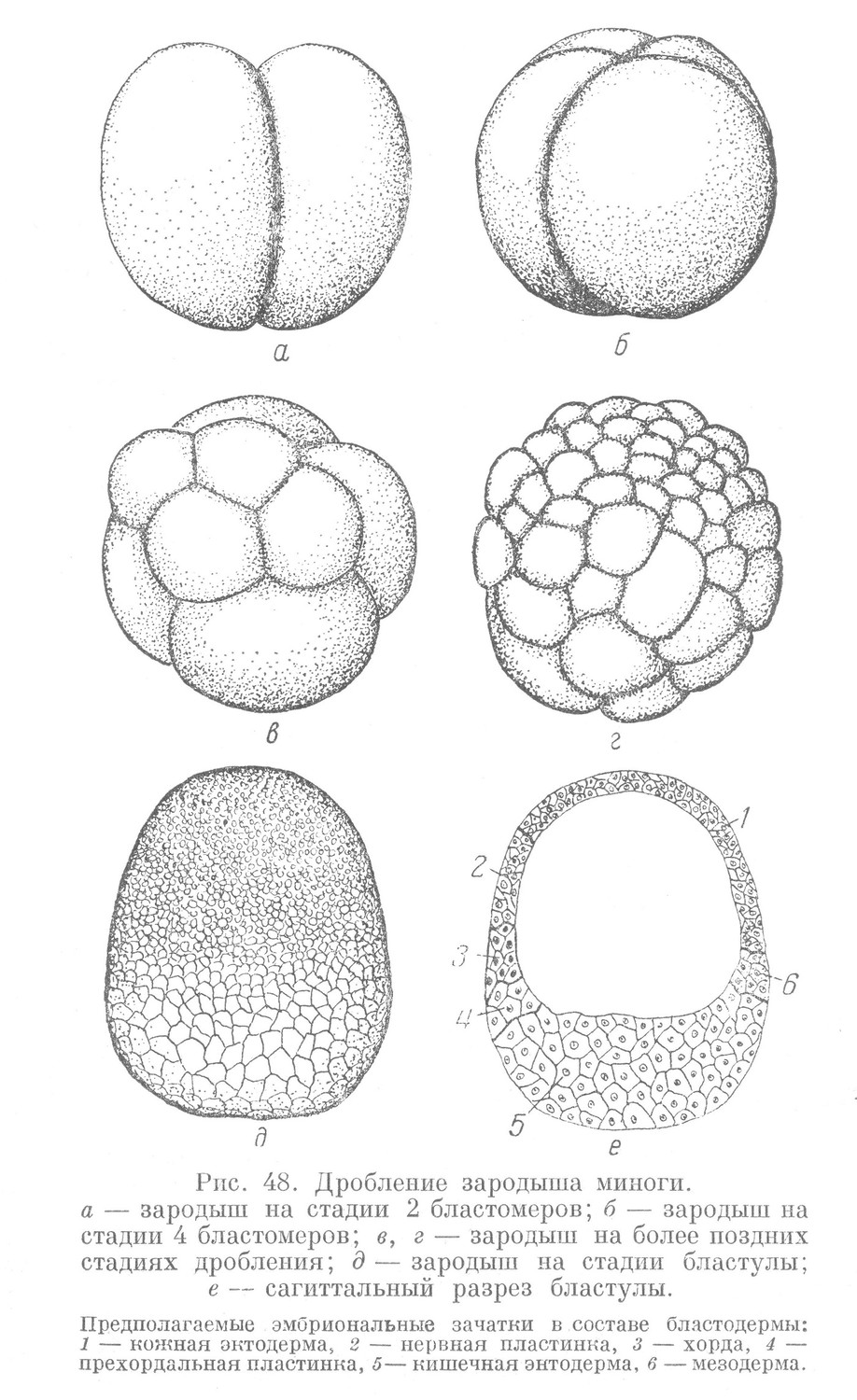
В ходе дробления наблюдается отставание деления бластомеров вегетативного полюса, содержащих желток, от мелких лишенных желтка бластомеров анимального полюса. В результате этого количество бластомеров вначале нарастает в геометрической прогрессии, а в дальнейшем этот порядок нарушается, и прогрессия представляет беспорядочно увеличивающийся ряд чисел (2, 4, 8, 16, 24, 32, 48, 64 и т. д.). Такое дробление называется полным, но неравномерным.
В ходе дробления, когда зародыш миноги состоит из 24—32 бластомеров, появляется тангенциальная борозда дробления. Она проходит в плоскости, параллельной наружной поверхности зародыша, разделяя каждый бластомер на два новых бластомера — наружный и внутренний. В результате чередования меридиональных, широтных и тангенциальных борозд дробления количество бластомеров в составе зародыша постепенно нарастает, а размер их постепенно уменьшается. В конце дробления зародыш имеет вид пузыря — бластулы, имеющей форму вытянутого эллипсоида (рис. 48, е). Бластодерма состоит из многих слоев клеток и в разных местах имеет неодинаковую толщину. Особенно толстой она представляется в области дна, где клетки содержат желток и вследствие этого имеют более крупную величину. Бластоцель заполнен жидкостью и располагается ближе к анимальному полюсу.
На основании данных изучения развития зародыша миноги с помощью метода маркировки бластодерму условно можно расчленить на участки, каждый из которых в период гаструляции является источником образования определенного эмбрионального зачатка. Эти предполагаемые зачатки расположены в бластодерме миноги так же, как и в бластуле ланцетника (рис. 47, а). Крышу бластулы образует материал будущих зачатков кожной эктодермы и нервной пластинки, а дно — материал кишечной энтодермы. Краевая зона образуется из материала будущих зачатков мезодермы, хорды и прехордальной пластинки.
Период гаструляции у миноги, так же как и у ланцетника, характеризуется двумя основными чертами. Вследствие более быстрого развития материала краевой зоны по сравнению с материалом крыши и особенно дна зародыш постепенно изменяет свою форму и из бластулы превращается в гаструлу. Внешне этот процесс проявляется в форме инвагинации, которая начинается у переднего конца зародыша серповидной бороздкой (рис 49,а).
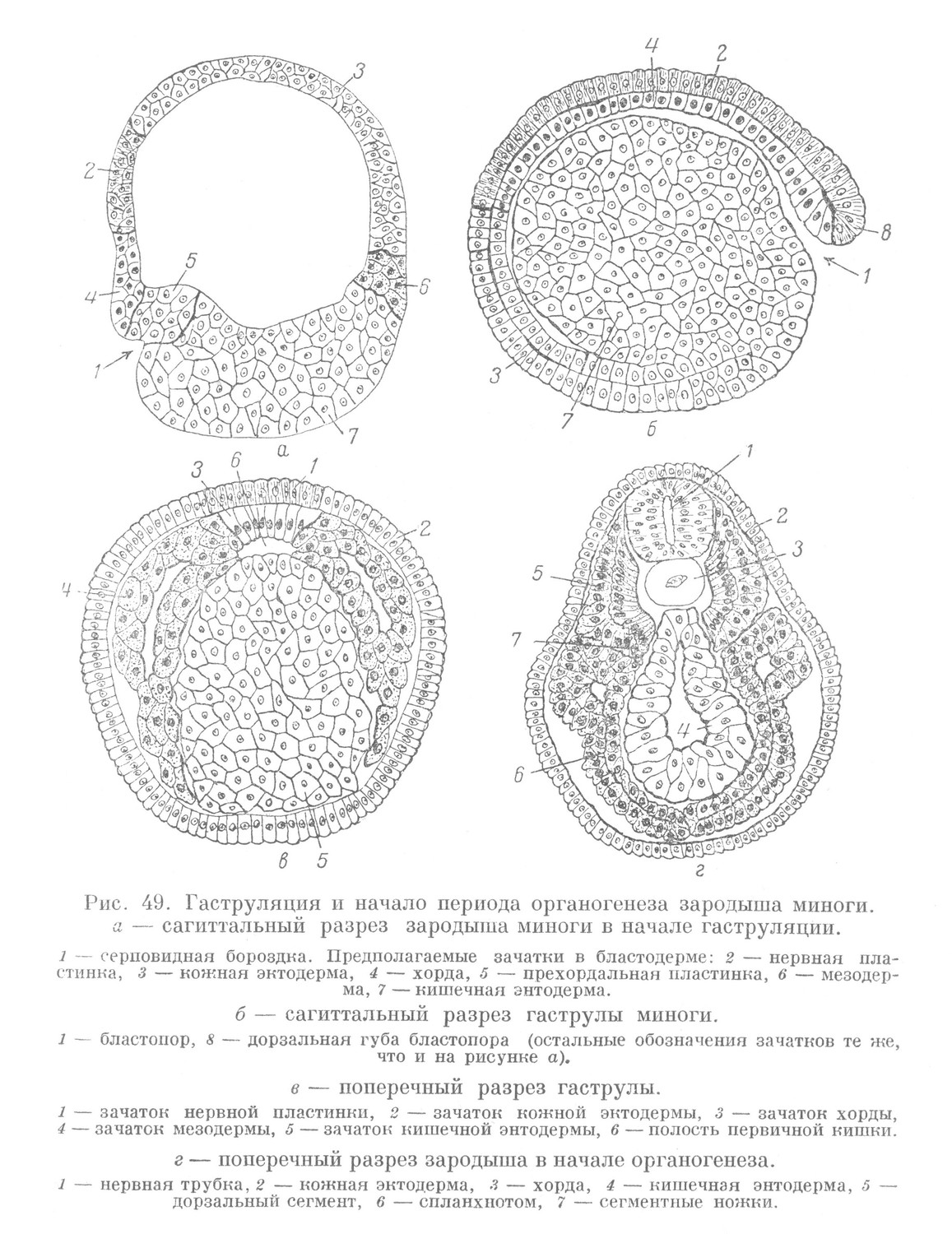
Последняя позже вследствие того, что материал дна погружается в сторону бластоцеля, углубляется. В гаструле миноги, так же как и у ланцетника, различается полость первичной кишки, или гастроцель, и ведущий в нее бластопор, ограниченный губами. В теле зародыша миноги выявляются различные эмбриональные зачатки, расположенные соответственно осевому размещению основных органов в теле позвоночных животных. Однако у зародыша миноги, в отличие от ланцетника, обособление зачатков начинается раньше, в конце гаструляции. Это проявляется тем, что значительная часть зачатка мезодермы из стенки первичной кишки выклинивается и распространяется на большое расстояние в щели между кожной эктодермой и кишечной энтодермой. Лишь небольшая часть ее находится по бокам от зачатка хорды, ограничивая просвет первичной кишки (рис. 49, в).
Что касается других зачатков, то они у миноги находятся в том же положении, как и в гаструле ланцетника. Начавшееся в ходе гаструляции обособление зачатков продолжается в периоде органогенеза. Происходит невруляция, в результате чего нервная пластинка, погружаясь под кожную эктодерму, превращается в нервную трубку. Следует отметить, что у миноги в процессе невруляции из наружного слоя гаструлы вместе с нервной трубкой выделяется зачаток, называемый ганглиозной пластинкой. Он образуется из материала, расположенного на стыке нервной пластинки с кожной эктодермой. При смыкании краев нервной трубки этот материал также сливается вначале в одну общую ганглиозную пластинку, которая располагается под замкнувшейся кожной эктодермой и над нервной трубкой. В ходе дальнейшего развития ганглиозная пластинка делится на правую и левую части, которые, расходясь в стороны, располагаются по бокам от нервной трубки. Каждая из них в свою очередь разделяется на отдельные участки, соответственно сегментам тела. Эти зачатки дают начало спинальным ганглиям, ганглиям вегетативной нервной системы и другим образованиям.
Из стенки первичной кишки выклинивается хорда и окончательно формируется мезодерма. Края кишечной энтодермы смыкаются, в результате чего возникает вторичная кишка (рис. 49, г).
Мезодерма разделяется на три тесно связанные между собой отдела. Одним из них являются дорзальные сегменты, каждый из которых соответствует определенному сегменту тела. Вентральный отдел мезодермы остается нерасчлененным, имеет вид мешка и называется спланхнотомом. Третьим отделом мезодермы являются сегментные ножки, составляющие вместе зачаток — нефротом. Они соединяют дорзальные сегменты со спланхнотомом. Последний состоит из двух листков. Один из них — внутренностный (висцеральный) прилегает к стенке вторичной кишки, другой — пристеночный (париетальный) находится в связи с кожной эктодермой.
В ходе органогенеза из мезодермы выделяются зачатки мезенхимы, которая очень рано начинает выполнять функцию ткани. Один зачаток мезенхимы выделяется в дорзолатеральном направлении от дорзальных сегментов, он называется кожной пластинкой, или дерматомом, другой зачаток выделяется в медио-вентральном направлении и называется склеротомом. Оставшиеся части дорзальных сегментов составляют зачаток, именуемый миотомом.
Висцеральный и париетальный листки спланхнотома также выделяют по зачатку мезенхимы: первый — в сторону кишечной энтодермы, второй — в сторону кожной эктодермы. Все 4 зачатка мезенхимы сливаются в один общий зачаток. Последний, разрастаясь между остальными зачатками тела зародыша, в ходе развития образует все разновидности тканей внутренней среды и гладкую мускулатуру. Миотом, выделившийся из дорзальных сегментов, дает скелетную мускулатуру, нефротом является источником развития выделительного, или почечного, эпителия, полость спланхнотома дает начало вторичной полости тела. Из оставшейся после выделения мезенхимы части листков спланхнотома образуется выстилка серозных полостей и другие родственные ей структуры.
Развитие рыб
Эмбриональное развитие различных рыб протекает неодинаково. Это зависит в значительной степени от количества желтка, находящегося в яйцевой клетке, и от других условий, в которых протекает развитие. В тех случаях, когда желтка в клетке относительно немного, развитие напоминает эмбриогенез миноги. Более резко отличается эмбриогенез у тех представителей класса рыб, яйцевая клетка которых содержит значительное количество желтка. Для этих видов характерно появление временных структур, которые следует рассматривать как признаки приспособления зародыша к окружающим условиям внешней среды.
Примером может служить развитие селяхий, яйцевая клетка которых является телолецитальной и содержит значительное количество желтка. Она достигает гигантских размеров. Так, яйцо акулы Lamna достигает 20 см и является самым крупным из яиц современных животных. Ядро располагается у анимального полюса яйца в участке цитоплазмы, содержащей сравнительно мало желтка. Строение зиготы остается неизученным.
Дробление у селяхий начинается появлением меридиональной борозды, которая разделяет зиготу лишь на небольшом протяжении у анимального полюса. Образуется стадия двух бластомеров, каждый из которых своим основанием переходит в общую нерасчлененную массу зиготы. Вторая меридиональная борозда проходит перпендикулярно первой и, так же как и первая, разделяет бластомеры лишь в области анимального полюса (рис. 50, а, б). Затем следует широтная борозда, которая соответствует экваториальной борозде зародышей ланцетника и миноги. В дальнейшем наблюдается чередование меридиональных, широтных и тангенциальных борозд, что приводит к образованию большого количества мелких бластомеров, образующих зародыш, имеющий форму дискобластулы. Образованием дискобластулы период дробления заканчивается (рис. 50, д, е).
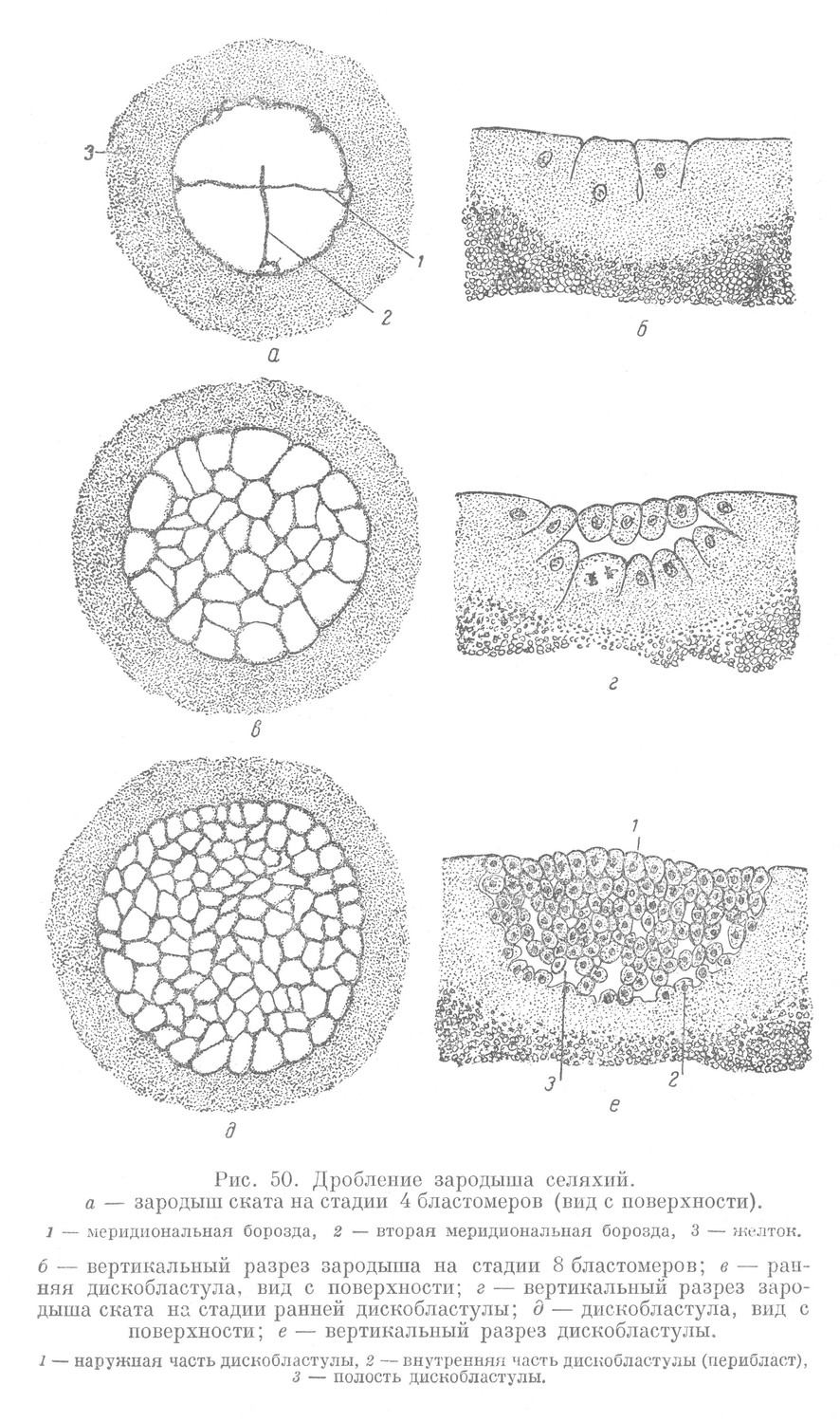
Этот тип дробления зародыша называется меробластическим или частичным. Такое дробление всегда завершается образованием дискобластулы, поэтому его называют также дискоидальным.
Дискобластула имеет стенку, состоящую из внутренней и наружной частей. Внутренняя часть стенки связана с нерасчлененным желтком и называется перибластом. Полость дискобластулы имеет вид сложной щели, которая заполнена жидкостью — продуктом жизнедеятельности клеток.
Особый интерес представляет наружная часть дискобластулы, так как здесь располагается материал, из которого в дальнейшем образуются окончательные органы тела зародыша (рис. 51, а).
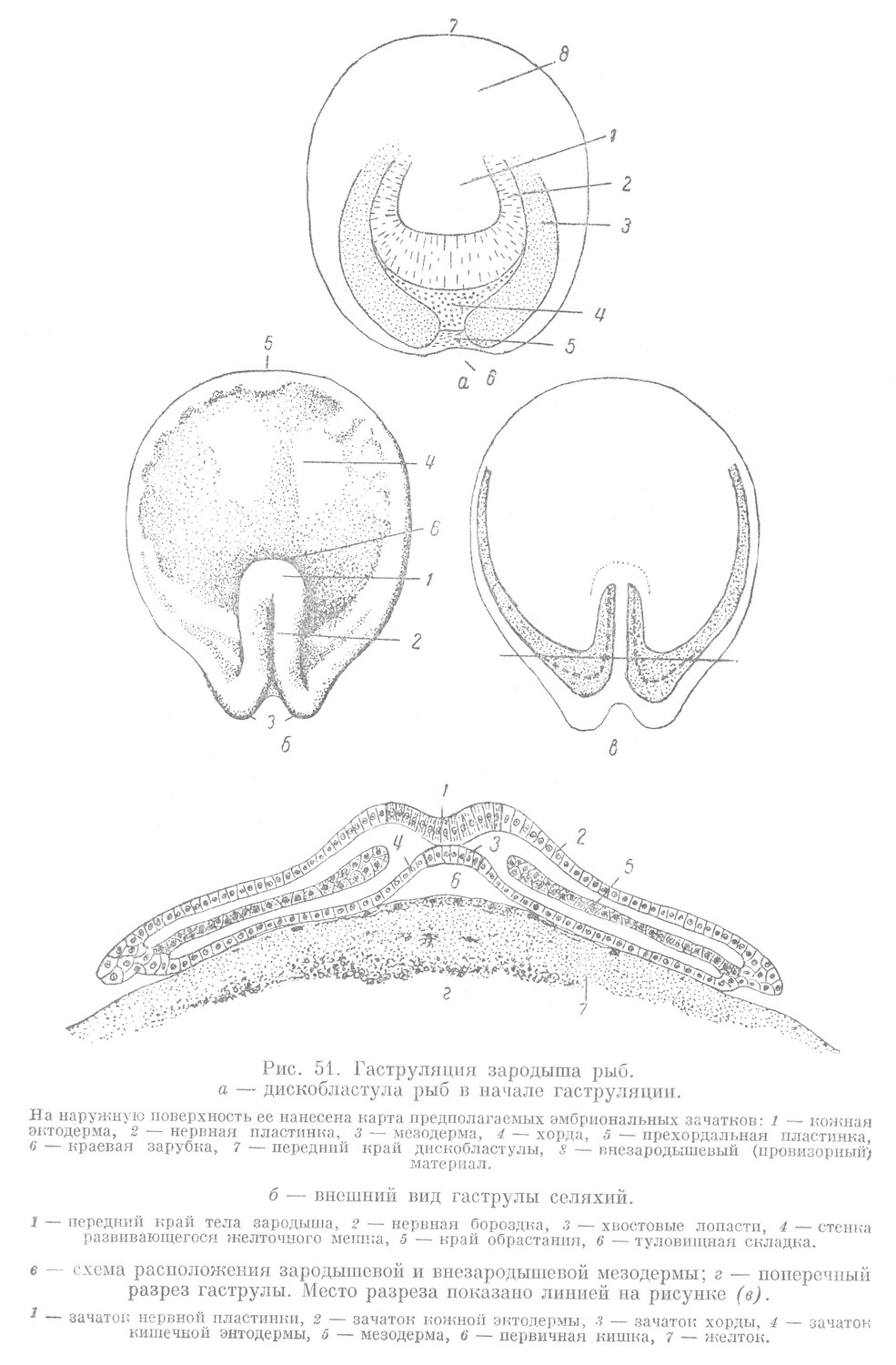
При рассматривании с поверхности дискобластула ската имеет вид широкого овала, имеющего передний и задний края, соответствующие переднему и заднему концам тела зародыша.
Опыты с маркировкой показывают, что материал, образующий дискобластулу, неоднороден. Значительная часть его дает начало развитию окончательных органов тела зародыша и поэтому называется зародышевым материалом. Кроме того, в составе дискобластулы содержится материал, являющийся источником развития временных, провизорных структур, связанных, главным образом, с функцией усвоения желтка как питательного материала. Этот материал называется внезародышевым, так как расположен вне тела зародыша и в его образовании непосредственного участия не принимает. Внезародышевый материал в дискобластуле находится как во внутренней его части, называемой перибластом, которая очень рано начинает резорбировать желток, так и у переднего края наружной части дискобластулы.
Метод маркировки дает возможность выявить в зародышевом материале дискобластулы отдельные участки, являющиеся источником развития зачатков будущих органов. Характер их расположения соответствует расположению, которое наблюдается в бластодерме ланцетника и миноги. Материал всех зачатков, за исключением зачатка кишечной энтодермы, находится в наружной части дискобластулы (рис. 51, а).
Рассмотрим развитие зародышевого материала и проследим, каким образом из него возникают отдельные оформленные эмбриональные зачатки и как образуется тело зародыша в целом. Неравномерное развитие отдельных участков зародышевого материала приводит к смещению их в составе зародыша в определенном направлении. Внешне это проявляется в том, что у заднего конца дискобластулы образуется небольшой вырез — краевая зарубка, которая по своему значению соответствует бластопору зародышей животных, рассмотренных нами выше. В связи с этим средняя часть зарубки будет соответствовать дорзальной губе, а склоны зарубки —- боковым губам бластопора. Это подтверждается тем, что через среднюю часть краевой зарубки смещается материал прехордалыюй пластинки и хорды, а через склоны краевой зарубки — материал мезодермы.
В конце гаструляции четко выявляются эмбриональные зачатки. Их расположение в теле зародыша соответствует принципу организации позвоночных животных (рис. 51, г).
Наиболее дорзально располагается зачаток нервной пластинки. С вентральной стороны к нему прилегает зачаток хорды. По бокам хорды находится зачаток мезодермы. Зачаток кишечной энтодермы лежит вентралънее от хорды и мезодермы.
Сравнивая взаимоотношения зачатков ската с аналогичной стадией у миноги, легко заметить общий принцип в расположении зачатков. У миноги, как уже указывалось, вычленение мезодермы из состава стенки первичной кишки начинается в конце периода гаструляции. У селяхий этот процесс идет значительно дальше. Здесь в конце гаструляции мезодерма полностью вычленилась из состава стенки первичной кишки. В головном отделе зародыша этот процесс пошел еще дальше, тогда как в хвостовом отделе он несколько запаздывает. Таким образом, у селяхий в конце гаструляции мезодерма уже вышла из стенки гастроцеля, в то время как у миноги небольшая часть ее еще находится в составе стенки (рис. 49, з и 51, г).
В период органогенеза диференцировка материала зачатков у селяхий продолжается: совершается невруляция, образуется вторичная кишка, мезодерма расчленяется на отдельные зачатки. Все это непосредственно предшествует развитию тканей из зачаткового материала.
Развитие внезародышевого материала происходит одновременно с развитием тела зародыша. Внезародышевый материал наружной части дискобластулы начинает быстро разрастаться по поверхности нерасчлененной желточной массы. Ввиду того, что зародыш находится в краевом положении, рост его в течение продолжительного времени идет эксцентрично (рис. 52, а).
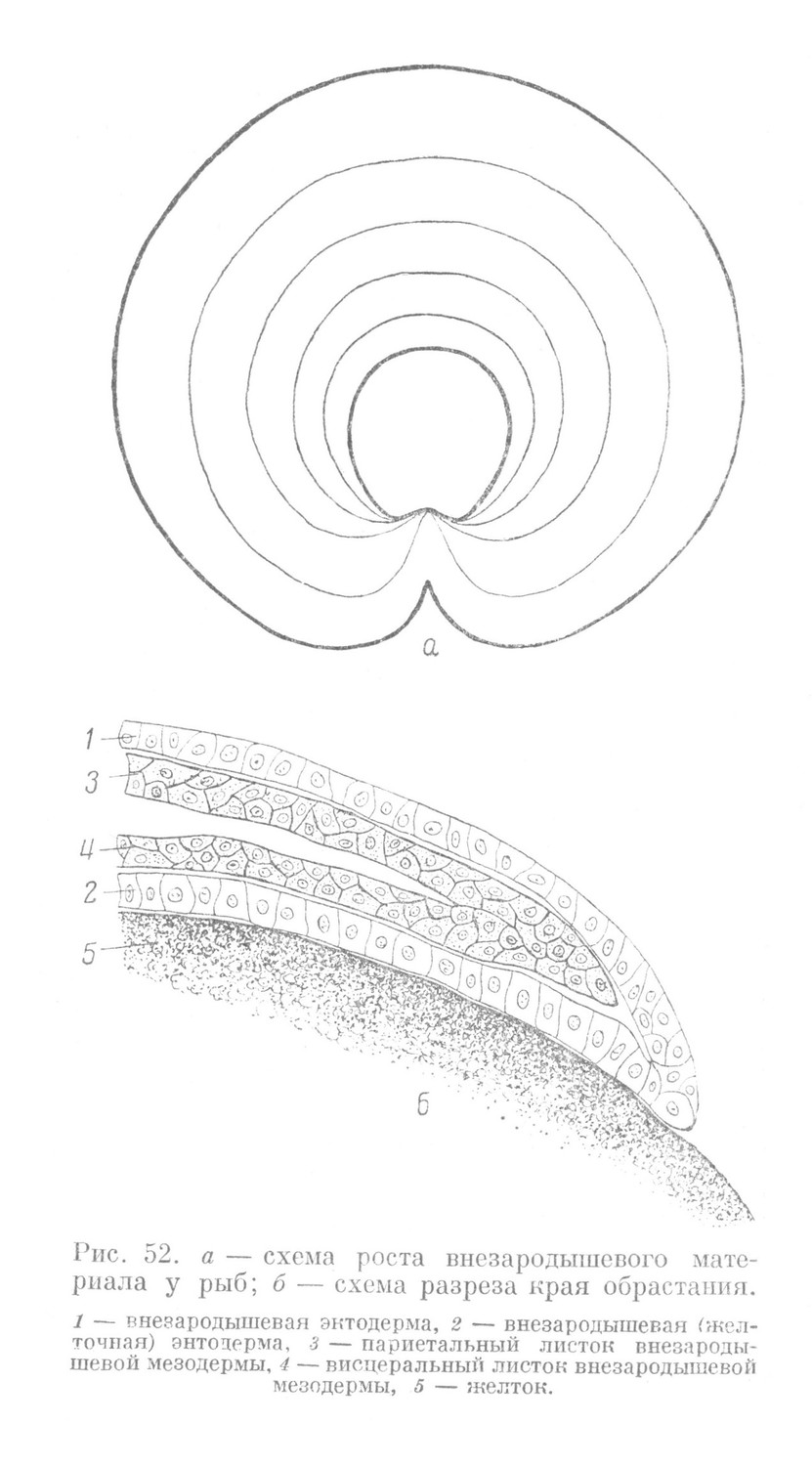
Край внезародышевого материала, продвигающийся по поверхности нерасчлененной желточной массы, называется краем обрастания (рис. 52, б). Он образован переходом внезародышевой эктодермы, составляющей наружный слой внезародышевого материала, во внезародышевую, или желточную, энтодерму. Последняя образует внутренний слой внезародышевого материала и прилегает непосредственно к желтку. Этот слой является продолжением перибласта, расположенного в области дискобластулы.
Между внезародышевой эктодермой и желточной энтодермой лежит внезародышевая мезодерма, разделенная на два листка. Один из них прилегает к внезародышевой эктодерме и называется париетальным, другой — связан с желточной энтодермой и называется висцеральным. У края обрастания оба листка мезодермы соединяются вместе, образуя край мезодермы.
После смыкания хвостовых лопастей развитие внезародышевого материала во всех направлениях происходит равномерно. Зародыш из краевого положения переходит в срединное. Однако окончательное смыкание краев внезародышевого материала происходит в соседстве с задним концом тела зародыша. Образование желточного мешка завершается полным смыканием краев роста.
Обособление тела зародыша от внезародышевого материала начинается постепенно в процессе гаструляции и в ходе развития желточного мешка. Это обособление связано с появлением туловищной складки, которая развивается в виде борозды, постепенно углубляющейся между зародышем и внезародышевым материалом (рис. 51, б). Такая борозда вначале появляется у переднего конца тела зародыша. Позднее она распространяется в каудальном направлении, отделяя боковые части зародыша и, наконец, замыкаясь на каудальном конце, отделяет эту часть тела зародыша от внезародышевого материала. В результате развития туловищной складки зародыш остается связанным со стенкой желточного мешка лишь тонким тяжем (желточным стебельком) (рис. 53). Последний представляет место перехода зародышевого материала в стенку желточного мешка. Здесь происходит переход кожной эктодермы во внезародышевую эктодерму, кишечной энтодермы — в желточную энтодерму и, наконец, зародышевой мезодермы (материала спланхнотома) — во внезародытевую мезодерму. Материал стенки желточного мешка, представленный указанными выше внезародышевыми зачатками, задолго до того, как окончательно сомкнутся края обрастания, начинает диференцироваться, превращаясь в ткани. Последние очень рано начинают функционировать. Впезародышевая эктодерма превращается в покровную ткань — эпителий. На месте желточной энтодермы возникает желточный эпителий. Внезародышевая мезодерма превращается в мезенхиму, которая, диференцируясь, дает начало кровеносным сосудам и крови. Таким образом, стенка желточного мешка очень рано получает тканевую организацию, представленную провизорными тканями (рис. 53 и 54).
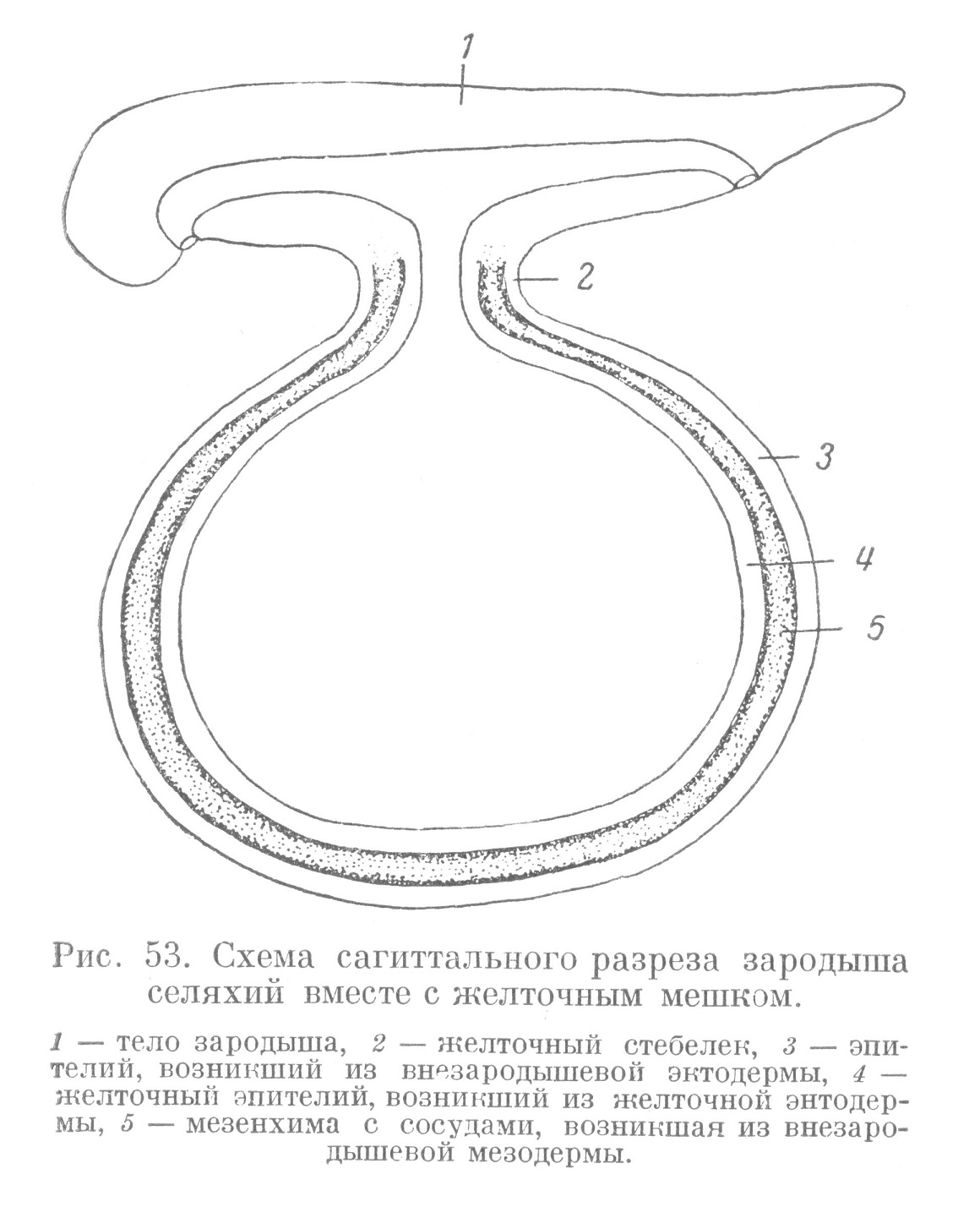
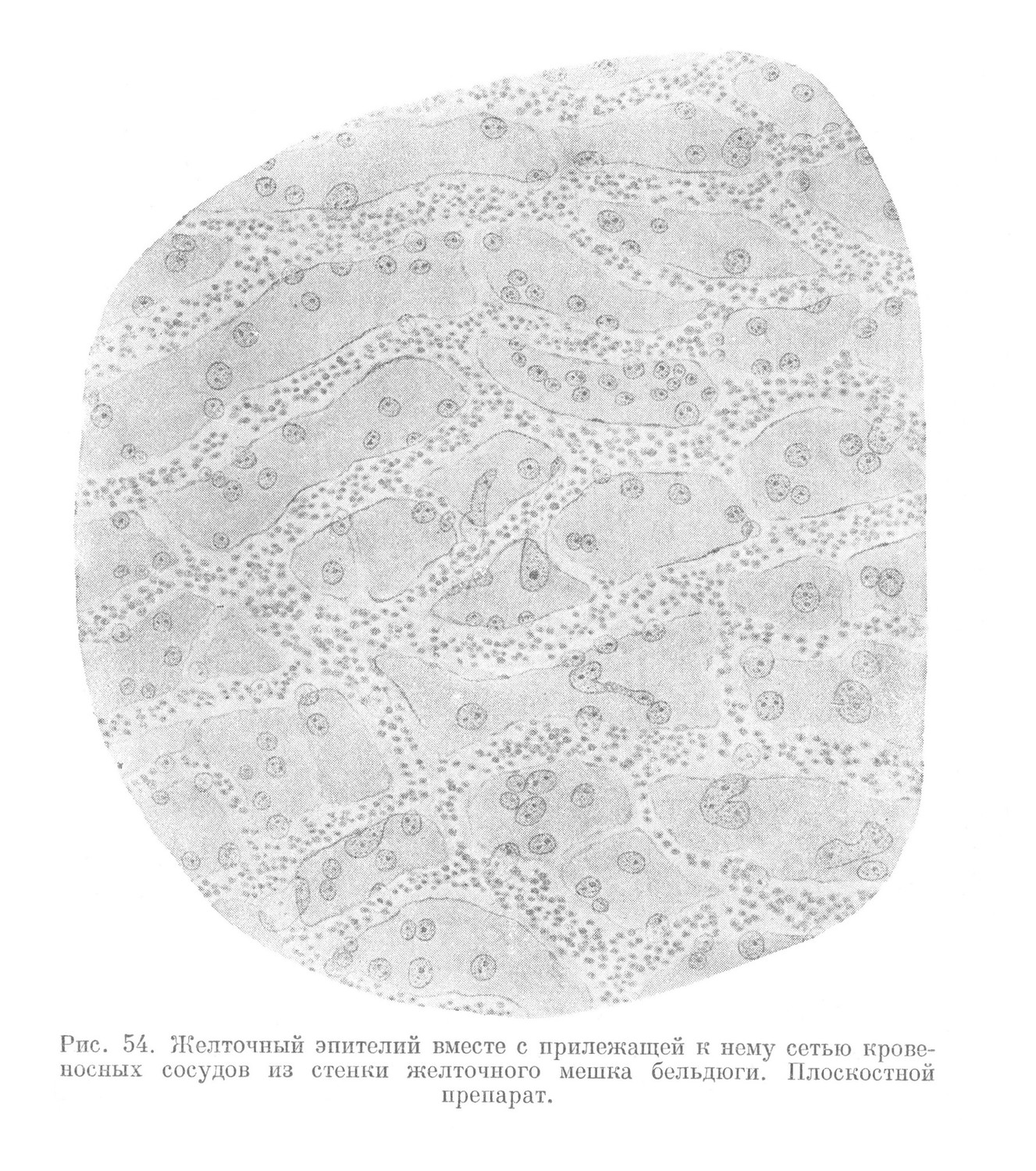
Основная функция желточного мешка — трофическая — осуществляется следующим образом. Желточный эпителий в процессе жизнедеятельности выделяет ферменты, разжижающие желток. Последний активно всасывается желточным эпителием. Далее питательный материал поступает в густую сеть кровеносных капилляров, которая тесно прилегает к основанию желточного эпителия (рис. 54). Отсюда питательные вещества по главным стволам желточных сосудов, проходящих в мезенхиме желточного стебелька, поступают в сосуды тела зародыша.
Описанное развитие зародыша селяхий отличается от голобластического типа развития, характерного для ланцетника и миноги, прежде всего тем, что в ходе его уже в конце дробления, наряду с зародышевым материалом, возникает внезародышевый. Такой тип развития называется меробластическим. Подобное развитие всегда сопровождается возникновением одного или нескольких провизорных органов. У рыб развивается лишь один провизорный орган трофической функции — желточный мешок.
Развитие амфибий
У амфибий, по сравнению с рыбами, наблюдается дальнейшее усложнение эмбриогенеза, хотя тип развития у большинства представителей этого класса является голобластическим. Рассмотрим для примера развитие лягушки и тритона.
Яйцевая клетка лягушки относится к телолецитальным яйцевым клеткам. Основная масса желтка в ней находится у вегетативного полюса. Поверхностный слой цитоплазмы на значительной поверхности яйца содержит бурый пигмент. Лишь в области вегетативного полюса яйцо лишено пигмента. После оплодотворения, которое совершается свободно в водной среде, в возникшей зиготе происходят изменения, напоминающие изменения в зиготе ланцетника. У лягушки они выступают менее отчетливо, ввиду наличия пигмента.
Дробление яйцевых клеток лягушки и тритона полное, но неравномерное. В конце дробления возникает бластула. Бластодерма у них многослойная, и в области дна она очень толстая, вследствие чего бластоцель располагается ближе к анимальному полюсу.
С помощью метода маркировки у лягушки и тритона удается проследить развитие отдельных участков бластодермы в зачатки органов, составляющих гаструлу. На основании этих данных, следуя ретроспективно от гаструлы к бластуле, бластодерму можно расчленить на отдельные, в определенном порядке расположенные, предполагаемые зачатки. Их расположение такое же, как и в дискобластуле рыб и в бластулах миноги и ланцетника (рис. 55, а).
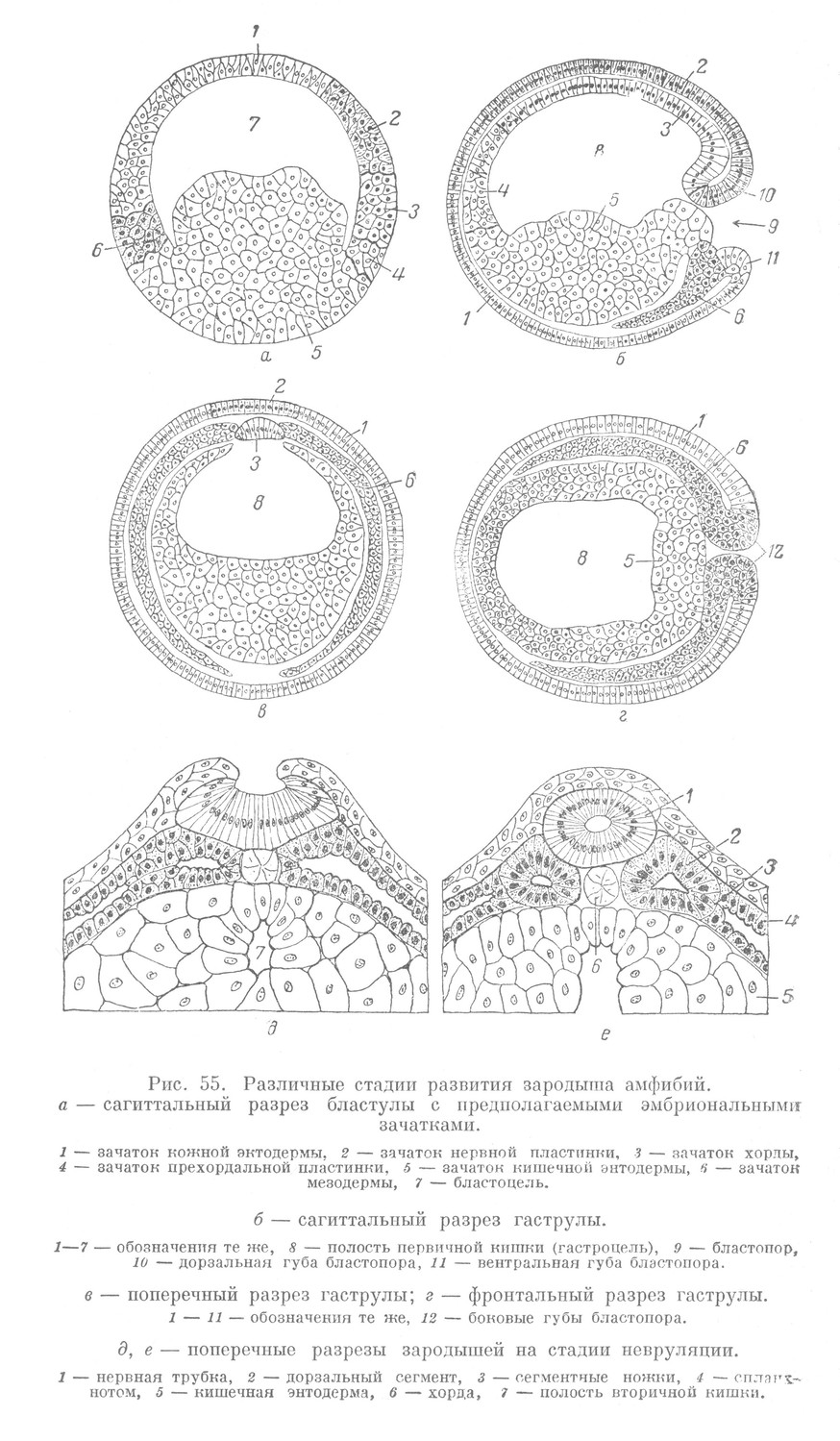
Период гаструляции у лягушки и тритона внешне протекает сходно с аналогичным периодом развития миноги. Он начинается появлением серповидной бороздки в области переднего конца зародыша. В дальнейшем происходит погружение материала дна бластулы в сторону бластоцеля. Зародыш постепенно изменяет свою форму, превращаясь в гаструлу (рис. 55, б, в).
Для этого периода развития характерны две вышеуказанные черты (неравномерный рост, смещение различных участков бластодермы и их качественное изменение). В конце гаструляции в теле зародыша отчетливо выявляются все основные зачатки, расположенные в определенном порядке, подобно аналогичной стадии развития у рыб, круглоротых и ланцетника. Однако выклинивание зачатков из состава стенки первичной кишки у амфибий начинается значительно раньше, чем у рыб.
Уже в начале гаструляции, в ходе образования первичной кишки, мезодерма вместе с хордой на значительном протяжении отделяется от кишечной энтодермы. В результате этого в развитой гаструле амфибий боковые и вентральная стенки первичной кишки образованы зачатком кишечной энтодермы, тогда как дорзальная стенка отсутствует (рис. 55,в). На ее месте имеется широкая щель, которая прикрывается хматериалом хордомезодермы, образующим как бы плащ. Последний располагается между кишечной энтодермой, с одной стороны, и нервной пластинкой и кожной эктодермой, с другой.
В период морфогенеза обособление эмбриональных зачатков продолжается (рис. 55, д, е). Совершается невруляция, в ходе которой возникает нервная трубка и ганглиозная пластинка. Края зачатка кишечной энтодермы смыкаются. Возникает вторичная кишка. Мезодерма расчленяется на дорзальные сегменты, сегментные ножки и спланхнотом. В дальнейшем из мезодермы выделяется зачаток мезенхимы.
Заканчивая рассмотрение основных черт эмбриогенеза низших позвоночных, следует отметить, что основным признаком при сравнительном изучении является характер расположения эмбриональных зачатков в теле зародыша в конце гаструляции. У всех рассмотренных животных, включая и ланцетника, зачатки органов тела зародыша в конце гаструляции располагаются однотипно. В этом проявляется палингенетический признак, указывающий на родственные отношения между всеми указанными позвоночными животными. Вместе с тем у рассмотренных выше животных наблюдается постепенное усложнение во взаимоотношении зачатков в период гаструляции. Если сопоставить поперечные разрезы гаструлы ланцетника, миноги, ската и лягушки, то станет ясно, что усложнение связано со все более ранним обособлением зачатков мезодермы, хорды и кишечной энтодермы, имеющих у зародышей перечисленных животных большее или меньшее отношение к первичной кишке (рас. 47, в, 49, в, 51, г, 55, в).
Что касается таких ценогенетических признаков, как, например, значительное содержание желтка в яйцевой клетке и появление в ходе развития желточного мешка, то эти факторы сами по себе еще не могут служить основанием, чтобы считать развитие рыб стоящим на более высокой ступени по сравнению с амфибиями, у которых и количество желтка в яйцевой клетке значительно меньше, и желточный мешок отсутствует. Напротив, основываясь на палингенетических признаках и, в частности, на характере взаимоотношений зачатков дефинитивных органов, следует прийти к противоположным и, несомненно, более правильным выводам и рассматривать эмбриогенез амфибий как дальнейшее усложнение по сравнению с эмбриогенезом рыб.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
|
