Переходя к рассмотрению основных черт развития млекопитающих, необходимо отметить, что у разных представителей этого класса эмбриогенез в деталях протекает различно. Среди млекопитающих имеется группа яйцекладущих (ехидна, утконос), развитие которых сходно с птицами.
У большинства млекопитающих развитие зародыша протекает в половых путях материнского организма и имеет место живорождение. Хотя последнее свойственно и отдельным представителям класса рыб, амфибий и рептилий, однако у них оно существенным образом отличается от живорождения у млекопитающих. В первом случае половые пути служат лишь местом для развития зародыша, который не имеет связи с телом матери и никаких питательных или других веществ не получает.
Живорождение у млекопитающих характеризуется тем, что зародыш получает необходимые для своего развития вещества из тела матери, что связано с развитием плаценты, соединяющей зародыш с тканями материнского организма.
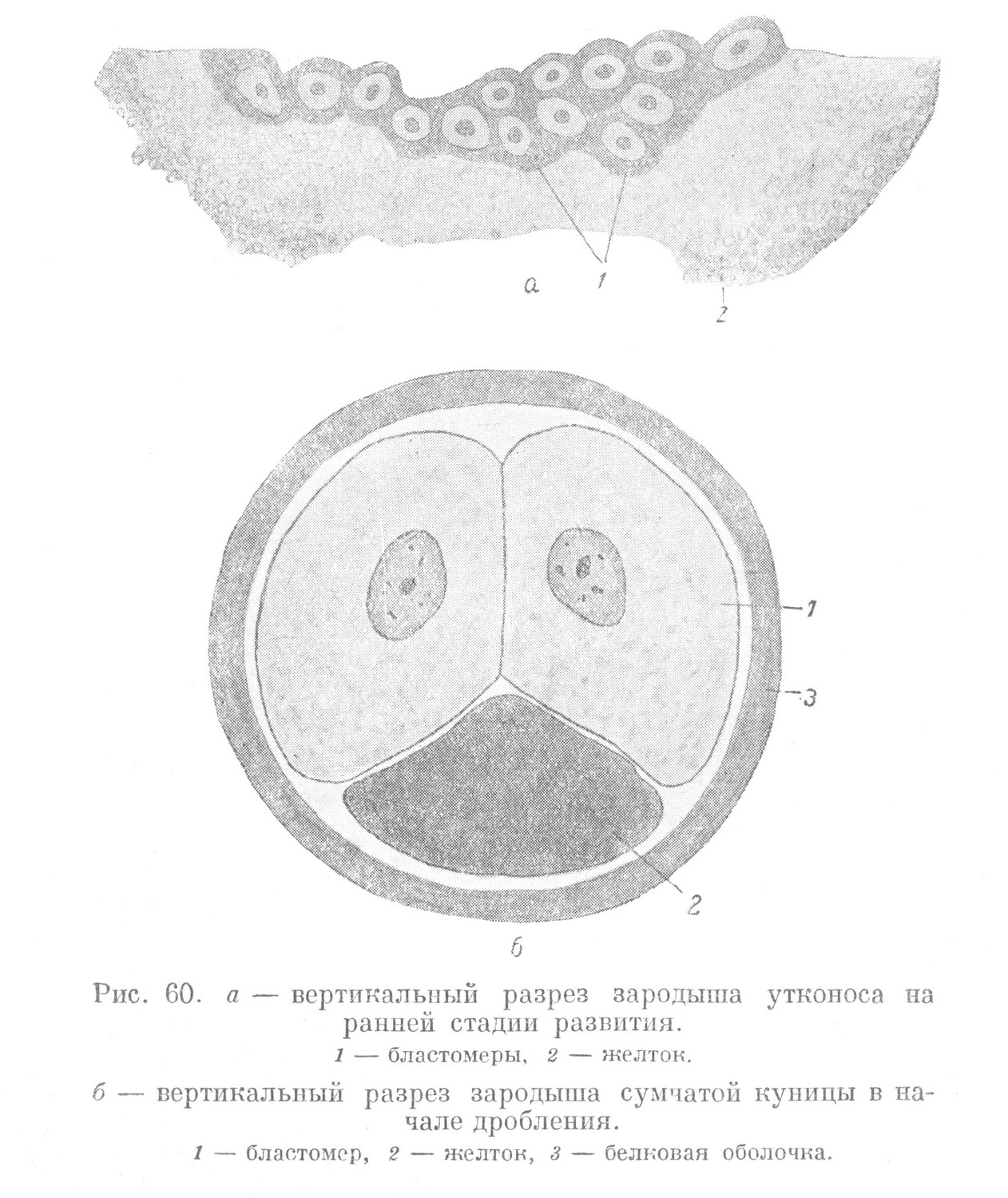
Яйцо яйцекладущих млекопитающих — телолецитальное, с огромным количеством желтка, поэтому оно имеет гигантскую величину. Развитие зародыша изучено слабо. Известно, что дробление яйца у этой группы млекопитающих, подобно птицам и рептилиям, частичное, меробластическое. Появляются те же и в том же порядке борозды дробления, что приводит к появлению многоклеточного зародыша на анимальном полюсе нерасчлененной желточной массы.
Из того, что известно о начальных стадиях развития яйцекладущих млекопитающих, представляет интерес раннее обособление бластомеров от желтка. Уже в начале дробления, после прохождения нескольких борозд дробления, все бластомеры отделены от желтка (рис. 60, а). У птиц полное отделение бластомеров от желтка происходит на более поздних стадиях дробления. У рыб этот процесс совершается значительно позднее, в конце дробления.
Из сопоставления приведенных фактов следует, что при меробластическом типе дробления у позвоночных полное отделение бластомеров от желтка происходит в разное время. У низших позвоночных это наблюдается в конце дробления, у высших — на более ранних этапах развития.
В связи с этими данными представляют интерес первые стадии развития сумчатых. Яйцевая клетка у них телолецитальная и имеет незначительные размеры. Небольшое количество желтка в ней располагается у вегетативного полюса. После оплодотворения начинается дробление. Первая борозда — меридиональная. Она начинается на анимальном и продолжается в сторону вегетативного полюса. Дойдя до той части зиготы, где располагается желток, борозда изменяет свое направление и становится широтной. В результате смыкания концов борозды зигота делится на два бластомера. Каждый из них полностью отделен от незначительного скопления желтка (рис. 60, б). Уже после прохождения первой борозды дробления, которую следует назвать меридионально-широтной, желток оказывается вне клеток. Из сказанного следует, что в ходе эволюции, при переходе от низших позвоночных к высшим, момент полного отделения бластомеров от желтка постепенно сдвигается к началу дробления. У сумчатых этот момент наблюдается при первом делении, когда появляется меридионально-широтная борозда. Следующие две борозды дробления у сумчатых — меридиональные. Образуется зародыш, бластомеры которого расположены на поверхности желтка в форме кольца. Та часть бластомеров, которая обращена к желтку, проявляет способность к его резорбции. Появляющаяся экваториальная борозда делит каждый бластомер на два новые, при этом один из них остается в связи с желтком. В ходе дальнейшего развития бластомеры, связанные с желтком, делятся быстрее и обрастают с поверхности скопление остальных бластомеров. Они не только резорбируют желток, но позднее и слой белка, образуя сплошной наружный слой зародыша, который называется трофобластом, так как выполняет трофическую функцию. Являясь начальной стадией развития провизорной временной структуры, он входит в дальнейшем в состав более сложного провизорного органа — хориона.
Таким образом, появление провизорной структуры у сумчатых происходит очень рано. Оно наблюдается после образования широтной борозды, которая возникает вслед за делениями в плоскости трех меридианов. Такое раннее выделение трофобласта и его быстрый рост объясняются чрезвычайно малым количеством питательного материала в зиготе. У зародыша очень рано возникает необходимость установления связи со стенкой половых путей матери, откуда в дальнейшем он и получает необходимые питательные вещества для своего развития.
Эмбриогенез млекопитающих будет рассмотрен на примере грызунов и хищников. В основу изложения развития этой группы животных положен эмбриогенез кролика, который изучен наиболее полно.
Развитие кролика
Яйцеклетка кролика — изолецитальная и содержит ничтожное количество желтка. Оплодотворение происходит в начальном отделе яйцевода. Строение зиготы изучено недостаточно. Дробление зародыша происходит в яйцеводе во время движения его в сторону матки. Первая борозда разделяет зиготу на два бластомера. Один из них более темный и имеет несколько меньшие размеры по сравнению с другим, более светлым бластомером (рис. 61, а). При следующем делении первым делится светлый бластомер, позднее — темный. В дальнейшем деление светлых бластомеров все более и более опережает деление темных. В результате увеличение количества бластомеров зародыша не укладывается в рамки геометрической прогрессии, а представляет ряд случайных цифр (2, 3, 4, 7, 10, 14 и т. д.). Такое деление называется полным и неравномерным, но отличным от полного и неравномерного дробления, которое имеет место у низших позвоночных, например у амфибий. Неравномерность дробления здесь связана с ускоренным развитием провизорного материала, представленного светлыми бластомерами, которые следует рассматривать как развивающийся трофобласт. Последний в составе зародыша занимает поверхностное положение. Обрастая скопление темных бластомеров, трофобласт очень рано устанавливает связь с материнским организмом. Исходя из сказанного о развитии светлых бластомеров и сопоставляя дробление у кролика с дроблением у сумчатых, следует, что первая борозда дробления у кролика является широтной, так как она аналогична широтной борозде у сумчатых, отделяющей первые бластомеры трофобласта.
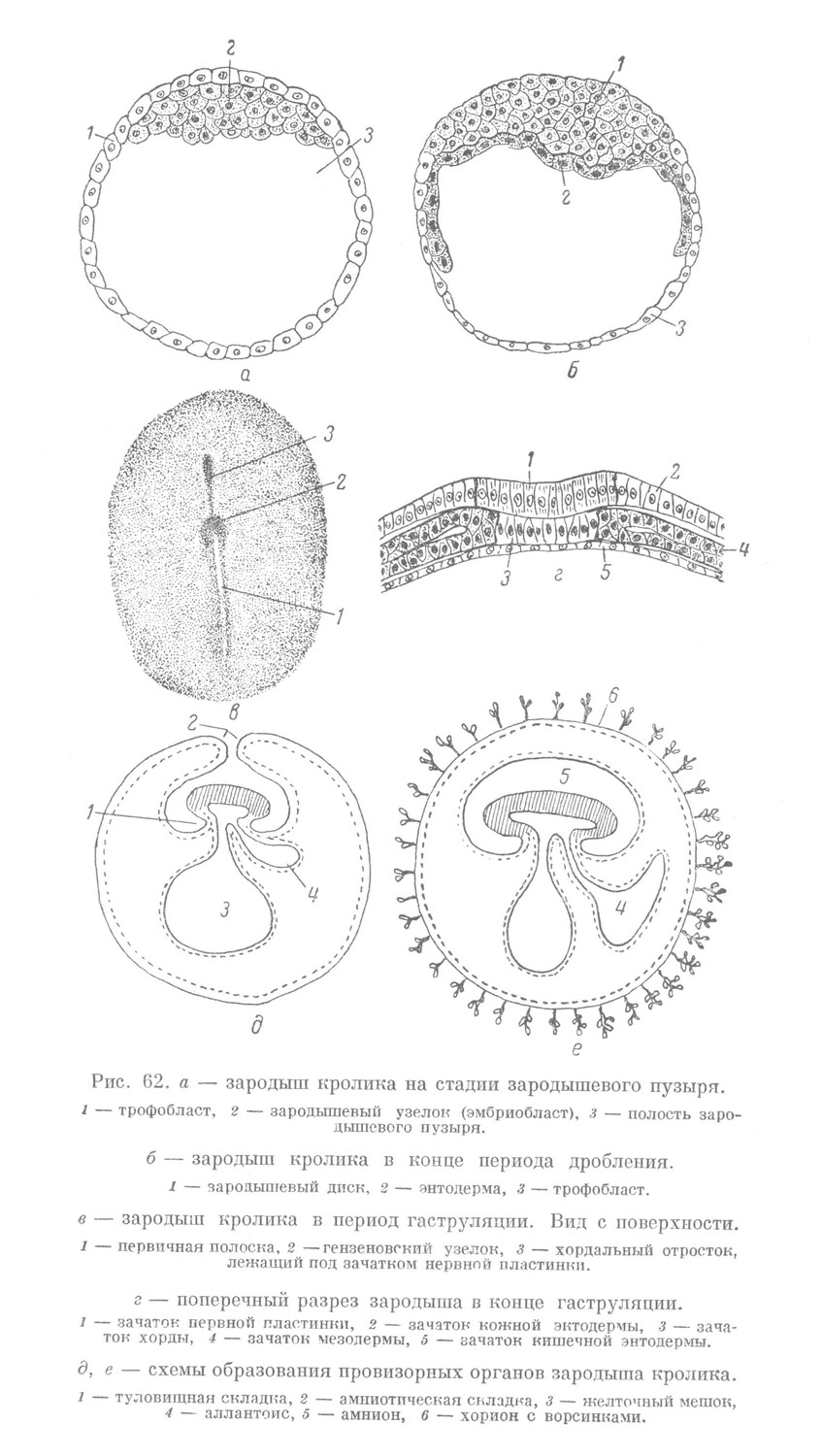
В полость матки зародыш попадает в виде плотного шара, называемого стерробластулой (рис. 61, д). В ней различают наружный слой — трофобласт и скопление более глубоко расположенных темных бластомеров — эмбриобласт. Эта стадия не является аналогичной бластуле или дискобластуле ранее рассмотренных позвоночных животных, так как отражает лишь одну из промежуточных стадий дробления, которое еще далеко от своего завершения. Такой зародыш, попадая в полость матки, вступает в связь со слизистой оболочкой, подготовленной к воспринятию зародыша.
С этого момента начинается имплантация — процесс внедрения зародыша в толщу слизистой оболочки и установление с ней органической связи. Имплантация протекает при активном участии трофобласта. Этот процесс будет рассмотрен ниже, при изложении развития и строения органа связи зародыша с материнским организмом.
В ходе имплантации период дробления завершается, при этом зародыш постепенно изменяет свое строение. В начале имплантации он из плотного шара превращается в зародышевый пузырь (рис. 61, е, 62, а). Стенка его образована тонким слоем трофобласта, состоящего из одного слоя клеток. Полость зародышевого пузыря постепенно увеличивается и достигает значительных размеров. Она заполнена белковой жидкостью, являющейся продуктом всасывания трофобласта.
Эмбриобласт, называемый зародышевым узелком, в одном месте связан с внутренней поверхностью трофобласта. В дальнейшем зародышевый узелок несколько распластывается и превращается в зародышевый диск, плотно прилегая к внутренней поверхности небольшого участка трофобласта. Затем этот участок трофобласта распадается, и зародышевый диск обнажается. Последний на всем протяжении смыкается своим краем с трофобластом, целиком входя в состав стенки зародышевого пузыря. Этой стадией завершается период дробления.
В зародышевом диске, который аналогичен дискобластуле, полость (бластоцель) отсутствует. Средняя часть его имеет небольшой размер и представляет зародышевый щиток. Вся остальная часть зародышевого диска образована внезародышевым материалом. В составе зародышевого щитка располагается материал зачатков органов, которые выявляются лишь в ходе гаструляции. Рассматривая развитие ретроспективно, от гаструлы к дискобластуле, зародышевый щиток можно условно расчленить на участки, соответствующие будущим зачаткам. Они располагаются в основном таким же образом, как и у зародышей круглоротых, рыб, амфибий и птиц.
Первым проявлением диференцировки внезародышевого материала является выделение внутреннего слоя зародышевого диска, обрастающего трофобласт со стороны полости. Этот материал, растущий в виде пласта, представляет энтодерму, которая на этой стадии развития еще не расчленяется на зародышевую и внезародышевую части (рис. 62, б). Разрастание ее напоминает развитие желточного мешка у рыб или птиц, поэтому этот процесс у млекопитающих рассматривают как образование желточного мешка. Отличие в данном случае состоит в том, что вместо желтка в мешке находится белковая жидкость. В дальнейшем начинается гаструляция.
В зародышевом щитке вследствие неравномерного роста происходит перемещение его отдельных участков. Подобно зародышу птиц и рептилии, здесь выявляется первичная полоска и гензеновский узелок (рис. 62, в). В ходе дальнейшей гаструляции от гензеновского узелка под наружным слоем в краниальном направлении вырастает головной или хордальный отросток. Последний представляет зачаток хорды. От первичной полоски по обеим сторонам хорды под наружным слоем вырастает зачаток мезодермы (рис. 62, г). В наружном слое над хордой выявляется в виде длинной полоски зачаток нервной пластинки, переходящий в зачаток кожной эктодермы. Вентрально от хорды и в связи с полостью зародыша находится зачаток кишечной энтодермы. За пределами зародышевого щитка этот зачаток переходит во внезародышевую энтодерму желточного мешка.
Таким образом, в ходе гаструляции в теле зародыша выявляются эмбриональные зачатки, расположенные соответственно общему плану организации тела позвоночных животных. Иными словами, здесь выявляется палингенетический признак, свойственный зародышам всех позвоночных животных. Далее происходит обособление тела зародыша от провизорного материала вследствие развития туловищной складки (рис. 62, д). Вначале она появляется в области головы и постепенно распространяется в каудальном направлении, отделяя в известной степени зародыш от вне- зародышевого материала.
Вместе с туловищной складкой появляется амниотическая складка, края которой, подобно складке у птиц, постепенно смыкаются над зародышем (рис. 62, д). Образуется амнион и аналогичный серозной оболочке птиц — хорион. Стенка амниона образована зачатками внезародышевой эктодермы и внезародышевой мезодермы.
У млекопитающих, так же как и у птиц, образуется аллантоис (рис. 62, д). Однако он никогда не достигает значительного развития. У приматов развитие его приостанавливается в самом начале. Он образуется, с одной стороны, из вентральной стенки задней кишки, с другой — из внезародышевой мезодермы. Упомянутые выше провизорные органы зародыша ускоренно развиваются, материал их диференцируется в ткани.
Желточный пузырь имеет значение как орган трофической функции лишь в самом начале. Эта функция выполняется им вместе с трофобластом. В дальнейшем трофическая функция полностью переходит к развивающемуся хориону.
Амнион достигает значительного развития и функционирует до конца эмбриогенеза. Материал стенки амниона быстро превращается в различные ткани. Пограничная ткань или эпителий амниона возникает из внезародышевой эктодермы. Так же как и у птиц, он представлен одним слоем плоских клеток. Вся остальная часть стенки амниона является производным мезенхимы, возникшей из внезародышевой мезодермы. Она представлена слоем малодиференцированной соединительной ткани, богатой клетками и имеющей небольшое количество промежуточного вещества в виде аргирофильных фибрилл и аморфного вещества. В этой ткани проходит сеть кровеносных сосудов. В связи с соединительной тканью находится слой гладкой мышечной ткани.
Наиболее сложное строение как в анатомическом, так и гистологическом отношении приобретает хорион (рис. 62, е). Образующий его трофобласт очень рано проявляет себя как функционирующая пограничная ткань. С другой стороны, в его образовании принимает участие внезародышевая мезодерма, дающая мезенхиму. Последняя образует соединительнотканную основу хориона, содержащую большое количество сосудов.
Хорион входит в состав плаценты или детского места — органа связи зародыша с материнским организмом. Так как строение плаценты весьма варьирует, то и хорион у разных животных неоднотипный. Хорион всегда обращен к тканям материнского организма, из которых он активно и избирательно всасывает необходимые для развития зародыша питательные вещества. Кроме того, через хорион осуществляются дыхательная и выделительная функции. Вместе с этим хорион выполняет еще одну важную функцию — защитную.
Хорион имеет очень сложное строение. На его свободной поверхности образуются разрастания, называемые ворсинками. Они в значительной степени увеличивают поверхность соприкосновения с тканями матери, что способствует выполнению трофической и других функций хориона.
Таким образом, у млекопитающих основными провизорными органами являются амнион и хорион. Последний объединяет несколько функций: трофическую, дыхательную, выделительную и защитную. Такое объединение различных функций в одном органе произошло в ходе эволюции как приспособление к условиям развития в теле матери. Зародыш, лишенный питательных веществ, пребывая с самого начала развития в матке, получает все необходимое для своего развития из окружающих тканей матери, с которыми он связан при помощи хориона. В силу этого развитие хориона начинается очень рано. Уже в результате первого деления выявляется светлый бластомер, дающий начало трофобласту. Последний вместе с мезенхимой образует в дальнейшем сложную структуру хориона. Более подробно развитие и строение названных провизорных органов будет рассмотрено ниже, при изложении эмбриогенеза человека.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 