Спинной мозг принимает чувствительные волокна задних корешков, являющиеся центральными отростками чувствительных клеток спинальных ганглиев, и содержит в своем составе конечные двигательные невроны, осевоцилиндрические отростки которых идут на периферию к мышцам в виде двигательных волокон передних корешков.
На поперечном разрезе спинной мозг представляется состоящим из двух симметричных половин, соединенных между собой узкой перемычкой, в которой проходит центральный спинномозговой канал (canal, centralis) (рис. 189). Последний является остатком полости нервной трубки. Спереди обе половины спинного мозга разделены глубокой передней щелью (fissura mediana anterior). В задней части спинного мозга щели нет; обе половины мозга сзади спаяны при помощи задней перегородки, которая имеет соединительнотканный характер (septum medianum posterior).
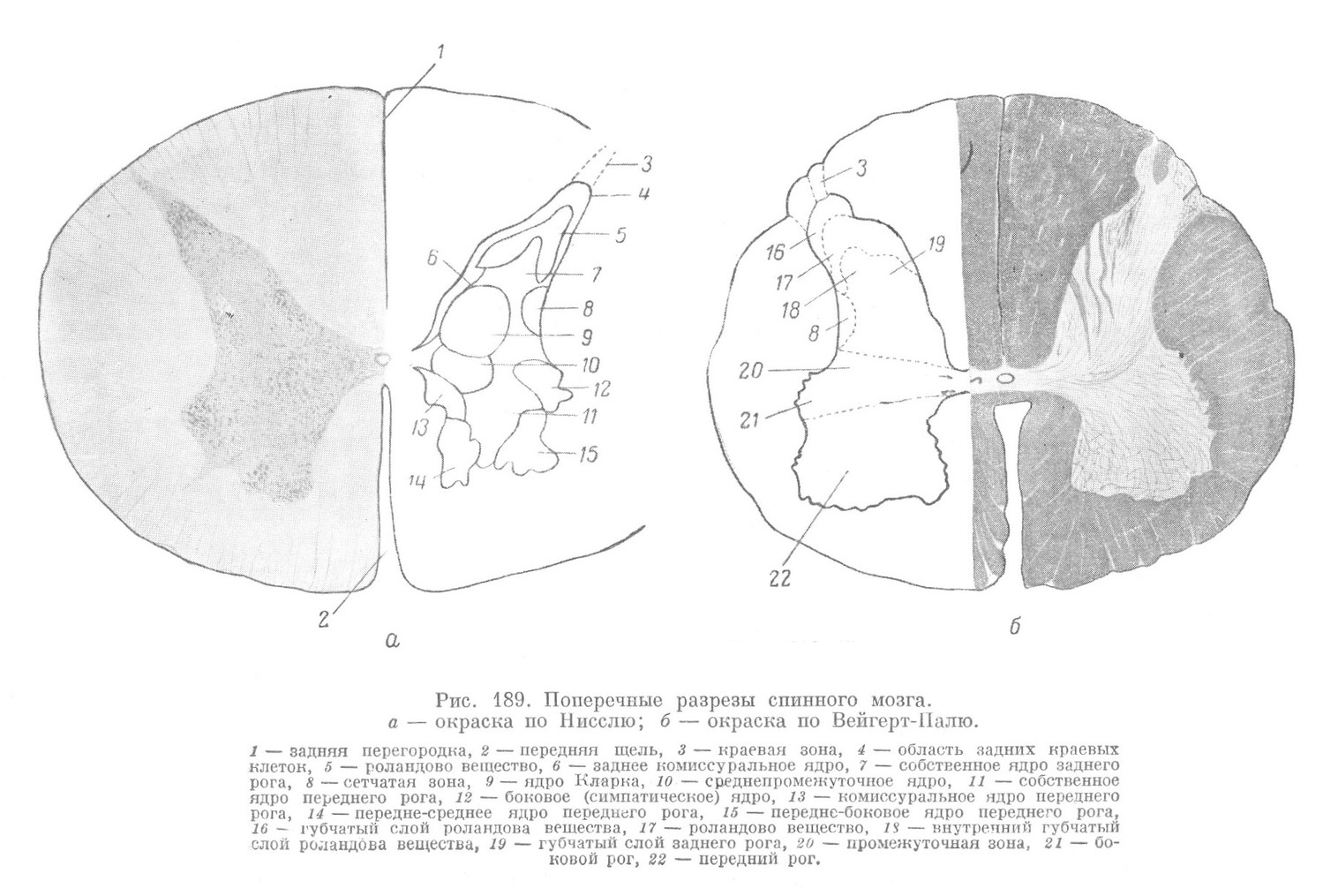
На поперечном разрезе спинного мозга уже невооруженным глазом удается заметить две отличающиеся своей окраской части: более светлую, лежащую снаружи, или так называемое белое вещество, и расположенную глубже темную массу — серое вещество (рис. 189, а). Серое вещество на поперечном разрезе имеет характерную форму — в виде буквы Н или бабочки.
Серое вещество мозга с каждой стороны образует два большие выступа по направлению к местам отхождения передних и вхождения задних корешков. Эти выступы называются рогами серого вещества. Различают передний, более объемистый рог серого вещества (cornu anterius) и более узкий — задний рог серого вещества (cornu posterius). Кроме этих двух главных рогов, серое вещество в некоторых отделах спинного мозга образует небольшой боковой выступ, носящий название бокового рога. Правая и левая массы серого вещества соединяются между собой перемычкой — серой спайкой (comissura grisea), в которой лежит центральный канал, разделяющий среднюю часть серой спайки на переднюю и заднюю серые спайки.
Кроме глиальных элементов, общих для всего спинного мозга, ceрoe вещество содержит в своем составе нервные клетки и волокна. Последние состоят главным образом, из безмякотных концевых разветвлений.
На поперечных разрезах серого вещества (рис. 189, б) можно отметить следующие нерезко отграниченные отделы (спереди назад): передний рог, промежуточную зону (pars intermedia) и задний рог. В последней кроме основной части, различаются губчатый слой (stratum spongiosum), переходящий сбоку в сетевидный отросток (formatio reticularis), вдающийся в белое вещество; роландово вещество (substantia gelatinosa Rolandi), ограниченное губчатой зоной (zona spongiosa) и переходящее в краевую зону Лиссауэра (zona marginalis Lissaueri). Такое разделение серого вещества основано, главным образом, на характере входящих в его состав волокон. Кроме волокон, в сером веществе имеются и нервные клетки. Они лежат определенными группами, которые получили общее название ядер. Картина расположения отдельных ядер представлена на рис. 189, а.
Белое вещество мозга в каждой его половине на поперечном разрезе подразделяется на 3 столба, отделенных друг от друга рогами серого вещества. Различают задние столбы белого вещества, расположенные между задней перегородкой и задними корешками, боковые,столбы, лежащие между передними и задними корешками, и передние столбы, ограниченные передней щелью и передними корешками. Впереди от серой спайки в средней перемычке имеется участок белого вещества, который соединяет между собой передние столбы и носит название белой спайки.
Белое вещество лишено нервных клеток и состоит преимущественно из продольно идущих мякотных нервных волокон. От серого вещества в белое вдаются радиально расположенные тонкие прослойки, состоящие преимущественно из глии. Они называются septula medullaria.
Соотношения белого и серого вещества, а равно и относительная мощность отдельных частей того и другого изменяются на всем протяжении спинного мозга. Так, в верхних отделах белого вещества больше, чем в нижних; передние рога серого вещества сильно развиты в шейном и поясничном утолщениях и т. д.
Все описанные анатомические структуры зависят от распределения глиальной стромы и нервных элементов, имеющих в спинном мозге вполне закономерное расположение. В своей совокупности они образуют ту систему, которая непосредственно управляет деятельностью органов движения, а спинной мозг содержит в своем составе аппарат, который тесно связан со всеми этими органами.
Аппарат спинного мозга млекопитающих и человека, имеющих сложно функционирующие конечности (особенно у человека, обладающего руками и высоко развитым головным мозгом), отличается значительной сложностью, что обусловлено филогенезом нервной системы. У примитивных организмов, лишенных конечностей и развитого головного мозга (круглоротые), спинной мозг имеет весьма примитивное строение, находящееся в полном соответствии с примитивными червеобразными движениями этих животных. На основе этого простого аппарата развился более сложный аппарат спинного мозга высших животных и человека.
В процессе эволюции усложнился и собственный аппарат спинного мозга и аппарат его связей с головным мозгом. При этом усложнении сохранились и все более примитивные, более древние механизмы. Последние в процессе филогенеза лишь оттеснились на задний план более новыми и более совершенными механизмами, развивавшимися одновременно с более совершенными органами движения и с более сложными связями с головным мозгом. Они соответствующим образом изменили свое функциональное значение.
Из сказанного следует, что одним из основных путей изучения спинного мозга является эволюционный метод. Понять строение спинного мозга можно, разделив его аппарат на группы, соответствующие тем этапам, через которые прошел спинной мозг в ходе своей эволюции.
Глиальный остов спинного мозга у низших позвоночных имеет чисто эпендимное строение. Центральный канал у них выстлан эпендимными клетками, разветвленные отростки которых, синцитиально связанные, доходят до поверхности мозга и образуют здесь пограничную глиальную перепонку. Такой глиальный остов в общем напоминает картины, какие обнаруживаются на известных стадиях развития спинного мозга у высших позвоночных животных.
У млекопитающих и человека преобладающую роль играет уже астроцитная макроглия, развившаяся на основе эпендимной глии, а последняя оказывается в значительной степени редуцированной. Эпендима выстилает центральный канал, и ее клетки дают лишь короткие и малоразветвленные отростки, доходящие до поверхности спинного мозга только в средней части — в области передней щели и задней септы (рис. 190). Вся остальная строма мозга построена из астррцитной макроглии, в которой при соответствующей обработке обнаруживаются длинно- и коротколучистые астроциты. Эта глия образует строму, имеющую различное строение в различных частях мозга. В сером веществе стромы имеются определенные территории, в которых лежат тела невронов. От местных особенностей глиального остова зависит выделение таких участков спинного мозга, как роландово вещество задних рогов и т. п. В белом веществе глиальный остов образует septula medullaria и все пограничные глиальные перепонки на поверхности мозга и вокруг кровеносных сосудов.

При описании нервных аппаратов спинного мозга их принято делить на 2 группы. В первую группу относят старый, основной механизм спинного мозга, который может быть назван собственным аппаратом или внутренним аппаратом спинного мозга. Ко второй группе относится аппарат двусторонних связей спинного мозга с головным.
Собственный аппарат спинного мозга обеспечивает известную способность этого отдела нервной системы к функционированию даже после отделения его от головного мозга. Невронные цепи собственного аппарата спинного мозга обеспечивают простейшие рефлексы. начинающиеся возбуждением чувствительной рецепторной точки на периферии и заключающиеся в переработке чувствительного импульса в двигательный, посылаемый в соответствующую мышцу.
Рефлекторные невронные цепи собственного аппарата обычно имеют трехчленный состав и слагаются из чувствительного. промежуточного, и двигательного невронов. Чувствительными невронами собственного аппарата являются клетки спинальных ганглиев (вероятно, мелкие и средней величины). Периферические их отростки, уходящие в спинномозговой нерв, заканчиваются в различных органах: (в частности, в мышцах) чувствительными рецепторными аппаратами. Центральные отростки входят в область краевой зоны задних рогов спинного мозга (рис. 191), где разветвляются на 2 веточки — одну восходящую, более длинную, другую нисходящую, более короткую которые идут вдоль спинного мозга в области краевой воны. Пройдя некоторое расстояние, каждая веточка загибается в серое вещество, где и заканчивается концевым разветвлением на теле одной из промежуточных, так называемых, пучковых клеток. Кроме того, и от восходящих, и от нисходящих колен центрального отростка чувствительного неврона в серое вещество отходят боковые веточки (коллатерали), также заканчивающиеся на телах пучковых клеток (из этих клеток на схеме рис. 191 изображена только одна).
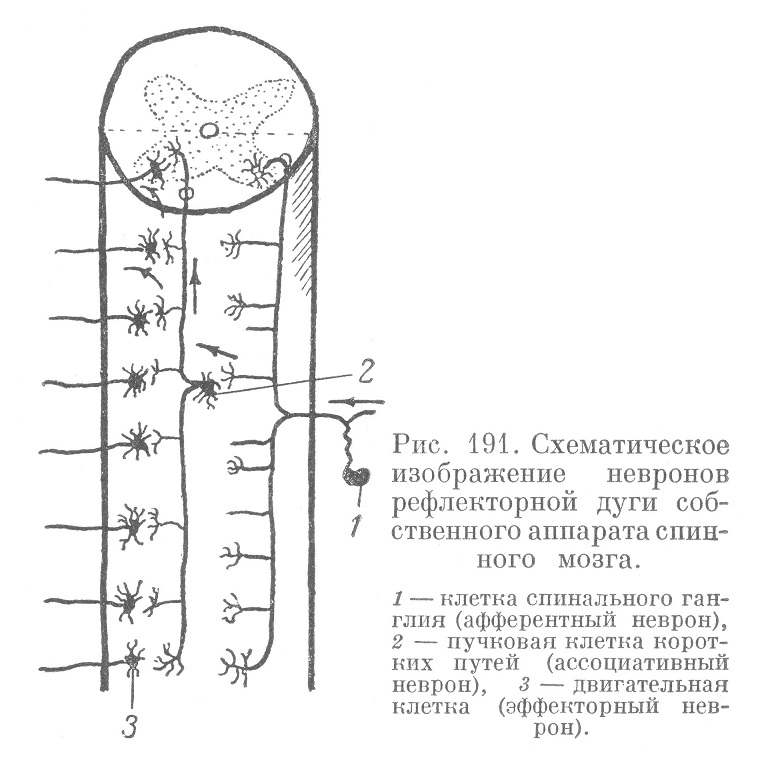
Пучковые клетки в спинном мозге лежат в задних рогах и в средней части серого вещества. По характеру отростков и их распространению пучковые клетки у высших позвоночных и человека делятся на несколько типов.
К собственному аппарату спинного мозга относятся только такие клетки, нервный отросток которых не выходит за пределы спинного мозга.
Это клетки средней величины, мультиполярные, с относительно короткими дендритами. Их нервный отросток проникает воелое вещество, где сразу по выходе из серого вещества разделяется (рис. 191) на более длинную, восходящую, и более короткую, нисходящую, веточки, идущие вдоль мозга в участках белого вещества, непосредственно прилегающих к серому. Обе эти веточки, так же как и отходящие от них коллатерали, заканчиваются в сером веществе, в его передних рогах, на телах расположенных там двигательных невронов (рис. 191).
Пучковые клетки собственного аппарата спинного мозга не одинаковы. Их нервный отросток может выходить в разные области белого вещества. В простейшем случае нервные отростки выходят в белое вещества бокового столба на той же стороне, где расположена и пучковая клетка (гомомерные клетки), нервный отросток других клеток выходит в белое вещество переднего столба. Имеются пучковые клетки, нервные отростки которых переходят на противоположную сторону в передние или боковые столбы белого вещества (гетеромерные клетки). Встречаются и такие пучковые клетки, нервный отросток которых вблизи клетки разветвляется на 2 веточки, из которых одна идет в белое вещество своей стороны, а другая переходит на противоположную сторону (гекатеромерные клетки).
Отростки пучковых клеток собственного аппарата мозга короткие и в белом веществе мозга прослеживаются в обе стороны на протяжении лишь 4—5 сегментов. Расположены они всегда в областях белого вещества, непосредственно прилежащих к серому, главным образом, в боковых и передних столбах (рис. 192).
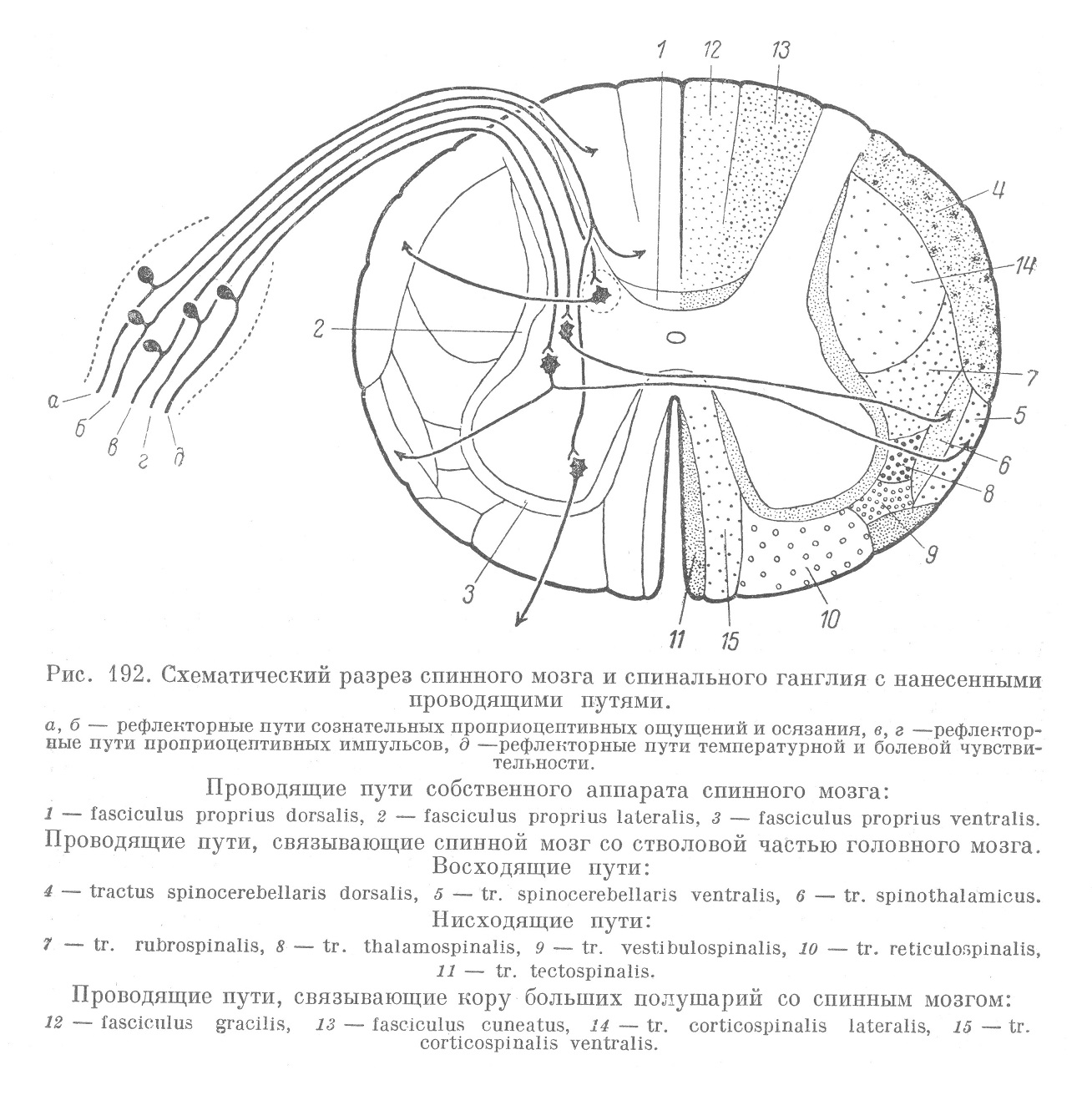
Таким образом, на протяжении всего спинного мозга серое вещество оказывается окруженным зоной белого вещества, содержащей только короткие внутренние проводящие пути спинного мозга, представленные отростками пучковых клеток собственного аппарата. Все эти пучки волокон называются собственными пучками спинного мозга. Рогами серого вещества, задней септой и передней щелью они делятся на 6 частей, вследствие чего в собственном проводящем аппарате в каждой половине мозга различают очень незначительный задний собственный пучок (funiculus proprius posterior), наиболее мощный боковой собственный пучок (fun. propr. lateralis) и передний собственный пучок (fun. propr. anterior).
Третьим невроном собственного аппарата спинного мозга является эффекторный двигательный неврон. Эти невроны наиболее крупные во всей нервной системе. Лежат они в передних рогах спинного мозга несколькими скоплениями, которые называются двигательными ядрами (рис. 189, а). У человека наибольшее число ядер содержится в передних рогах шейного утолщения спинного мозга, откуда иннервируются верхние конечности.
Двигательные клетки мультиполярны и имеют относительно длинные дендриты, лежащие в сером веществе спинного мозга. Их нервный отросток выходит непосредственно в передний корешок и дает начало двигательному волокну, заканчивающемуся в мышцах эффекторным двигательным концевым аппаратом.
Таким образом, в состав собственного аппарата спинного мозга входят:
1) чувствительные клетки спинальных ганглиев с короткими центральными отростками
2) пучковые клетки с короткими невритами, рассеянные в сером веществе задних рогов и промежуточной области,
3) двигательные клетки, лежащие в сером веществе передних рогов и образующие своими нервными отростками передние корешки. Проводящими путями собственного аппарата в белом веществе являются передние, боковые и задние собственные пучки, содержащие отростки пучковых клеток, и так называемая терминальная зона (рис. 189, а, б), составленная центральными отростками чувствительных клеток собственного аппарата.
Всякое возбуждение чувствительного неврона распространяется на целый ряд промежуточных пучковых клеток (рис. 191), а через эти последние — на еще большее число двигательных эффекторных невронов. Происходит, как говорят, «лавинообразное нарастание рефлекса». Следовательно, из одной точки можно возбудить значительную эффекторную область. У низших позвоночных, совершающих рыбообразные движения всем туловищем, собственный аппарат в спинном мозге распределяется своими элементами достаточно диффузно. У высших животных и у человека он значительно более диференцирован, особенно в тех местах, которые связаны с иннервацией конечностей. Здесь двигательные невроны собраны в виде отдельных ядер, из которых каждое имеет определенную область иннервации и определенные связи в самом внутреннем аппарате. Трехчленный рефлекторный аппарат спинного мозга является аппаратом филогенетически очень древним.
У высших позвоночных, кроме трехчленных рефлекторных дуг, имеются и двучленные. В этом случае коллатерали центральных отростков чувствительных клеток оканчиваются непосредственно на двигательных невронах передних рогов. В таком аппарате возбуждение передается менее распространенно, более точно, так как возбуждается меньшее количество эффекторных невронов. Аппарат прямых чувствительно-двигательных связей является аппаратом филогенетически более новым.
Собственный аппарат спинного мозга имеет двусторонние связи с аппаратом головного мозга, который объединяет работу всей нервной системы.
Головной мозг представляет образование весьма сложное, претерпевшее длительную эволюцию. В головном мозге следует различать заднюю (бульбарную) часть, представленную продолговатым мозгом, который у низших форм, наряду со средним мозгом и основанием большого мозга, играет доминирующую роль. С этой частью связана иннервация полости рта, которая осуществляется рядом черепных нервов, и органа равновесия (слуха). В бульбарной части у низших позвоночных сосредоточены также центры, объединяющие работу спинного мозга.
Передний отдел головного мозга, представленный средним (mes-), промежуточным (di-) и конечным (telencephalon) мозгом, связан, главным образом, с основными органами чувств — зрения и обоняния, определяющими поведение всех животных. В большом мозге низших позвоночных развито только основание (полосатые тела), непосредственно связанное» (так же, как и основание промежуточного мозга — чертоги, или зрительные бугры) с органом обоняния; средний мозг этих животных является главным зрительным центром.
В процессе эволюции, параллельно с диференцировкой головного мозга, между ним и спинным мозгом устанавливались все более и более диференцированные связи. Вначале эти связи осуществлялись через бульбарный отдел головного мозга, где заканчиваются пути, приносящие импульсы из спинного мозга и из передних отделов головного мозга, и где начинаются центробежные координирующие пути к спинному мозгу.
В дальнейшем в спинном мозге установились непосредственные двусторонние связи со все более и более развивающимися передними частями мозга: с вестибулярным аппаратом (органом равновесия), со средним мозгом и, наконец, со зрительным бугром (thalamus). У млекопитающих, особенно у человека, мощное развитие получает новый аппарат головного мозга, который является надстройкой над всей нервной системой; это — кора большого мозга.
У млекопитающих и у человека в связи с этим появляется и новый аппарат связей со спинным мозгом. Устанавливается двусторонняя связь собственного аппарата спинного мозга с корой большого мозга. В процессе эволюции более древние общие связи уступают место более диференцированным новым, и последние начинают играть преобладающую роль, суживая значение старых общих связей.
Однако и старые связи из системы не исчезают, а сохраняются, занимая в ней подчиненное положение. Таким образом, и у млекопитающих, и у человека спинной мозг имеет связи и со стволовой частью головного мозга, начиная с бульбарного ее отдела и кончая зрительными буграми, и с корой большого мозга.
В аппарате спинного мозга связи со стволовыми частями головного мозга (флогенетически более старые) и связи с корой (новейшие) осуществляются различно.
Первые образованы пучковыми клетками с длинными нервными отростками, идущими в восходящем направлении. Такие пучковые клетки часто располагаются определенными группами или ядрами и в задних рогах и в средних частях серого вещества. На телах этих мультиполярных клеток заканчиваются коллатерали центральных отростков чувствительных клеток спинальных ганглиев, а нервные их отростки направляются в белое вещество на одноименную или противоположную сторону; здесь они загибаются кверху и направляются в ствол головного мозга, где и заканчиваются в той или иной его части. Все волокна этих восходящих связей в спинном мозге сосредоточены в поверхностных частях боковых столбов белого вещества.
В боковых столбах у человека и млекопитающих различают так называемый передне-боковой пучок (рис. 192, 5 и б) и дорзальный пучок (рис. 192, 4). Дорзальный пучок содержит волокна, идущие к мозжечку, почему он и называется дорзальным (прямым) мозжечковым путем (tractus spinocerebellaris dorsalis). Пучковые клетки, дающие начало волокнам этого пути, сосредоточены в задних рогах на одноименной стороне в виде хорошо выраженных ядер, называемых ядрами Кларка (рис. 189, а, 192). Передне-боковой пучок составляется из волокон, являющихся нервными отростками пучковых клеток, расположенных в основании заднего рога противоположной стороны и образующих так называемое собственное ядро заднего рога (рис. 189, а, 192).
В поверхностных частях передне-бокового пучка сосредоточены волокна, идущие к мозжечку и образующие так называемый вентральны (перекрещенный) путь к мозжечку (tr. spinocerebellaris ventralis). В более глубоких частях передне-бокового пучка располагаются волокна, идущие к зрительному бугру; они образуют спиноталамический путь (tr. spinothalamicus).
В передне-боковом пучке сосредоточены и незначительно развитые у человека пути к продолговатому и среднему мозгу. В шейной области из этих путей выделяется пучок, идущий к оливам (tr. spinoolivaris) и обеспечивающий связь с вестибулярным аппаратом. Этот пучок имеет важное значение, так как при его помощи осуществляется координация движений туловища и головы, в которой лежит орган равновесия.
В переднем отделе боковых столбов, между передне-боковыми и собственными пучками, а отчасти в передних столбах белого вещества, лежат все нисходящие пути, идущие из ствола головного мозга и соответствующие восходящим связям передне-бокового пучка. Здесь лежат пути, начинающиеся от клеточных ядер промежуточного, среднего и продолговатого мозга. Среди этих путей следует отметить двигательный путь, идущий из красного ядра (tr. rubrospinalis), и путь, идущий из вестибулярного ядра (tr. vestibulospinalis) и из бульбарной части (tr. reticulospinalis). Здесь же проходит и путь из зрительного бугра (tr. thalamospinalis) (рис. 192).
Волокна, составляющие все эти обратные нисходящие пути, оканчиваются терминальными коллатералями на клетках двигательных ядер передних рогов (рис. 193). Характерной особенностью всех этих нисходящих путей является то, что они от места своего начала и до двигательного ядра переднего рога идут не прерываясь.
Некоторое представление об описанных связях спинного мозга и отношении к ним его собственного аппарата можно получить из схематических рисунков 193 и 194. На этих рисунках представлены разрезы спинного мозга и основных отделов головного мозга. В переднем роге спинного мозга (рис. 193) изображена одна большая двигательная клетка, олицетворяющая собой первичный двигательный центр. В соответствующем этому сегменту спинного мозга спинальном ганглии изображены 5 чувствительных клеток. Два афферентных неврона, отростки которых представлены тонкими линиями, относятся к собственному аппарату спинного мозга. Один из них связан с пучковой клеткой коротких путей и относится к трехчленной рефлекторной дуге. Другой устанавливает связь непосредственно с двигательной клеткой и входит в состав двухчленной рефлекторной дуги. Афферентный неврон, отростки которого изображены толстым пунктиром, в основании заднего рога передает импульс пучковой клетке длинных путей. Это является началом перекрещенного пути к зрительному бугру (tr. spinothalamicus) (рис. 194), откуда нисходящее волокно (обозначенное также пунктиром) направляется к двигательной клетке переднего рога (tr. thalamospinalis). Клетка спинального ганглия, отростки которой обозначены толстой поперечно исчерченной линией, передает импульс пучковой клетке длинных путей, лежащей в составе ядра Кларка. Она является началом сложного рефлекторного пути (tr. spinocerebellaris dorsalis), идущего через мозжечок (рис. 194), где в двух центрах происходит переключение импульса. После этого проводник этого рефлекторного пути направляется (в составе ножек мозжечка к четверохолмию) к красному ядру среднего мозга, где и происходит передача импульса следующему неврону. Аксон последнего и входит в состав нисходящего пути (tr. rubrospinalis), передающего импульс двигательной клетке переднего рога (рис. 193). Подобным же образом осуществляются двусторонние связи и с другими отделами ствола головного мозга. Рефлекторный путь, идущий от вестибулярного аппарата внутреннего уха к ядрам вестибулярного нерва, лежащим в области моста, и нисходящий от ядер путь (tr. vestibulospinalis), передающий импульс двигательной клетке переднего рога, обозначены на рис. 193 и 194 толстой, косо исчерченной линией. Через этот рефлекторный путь осуществляется равновесие тела.
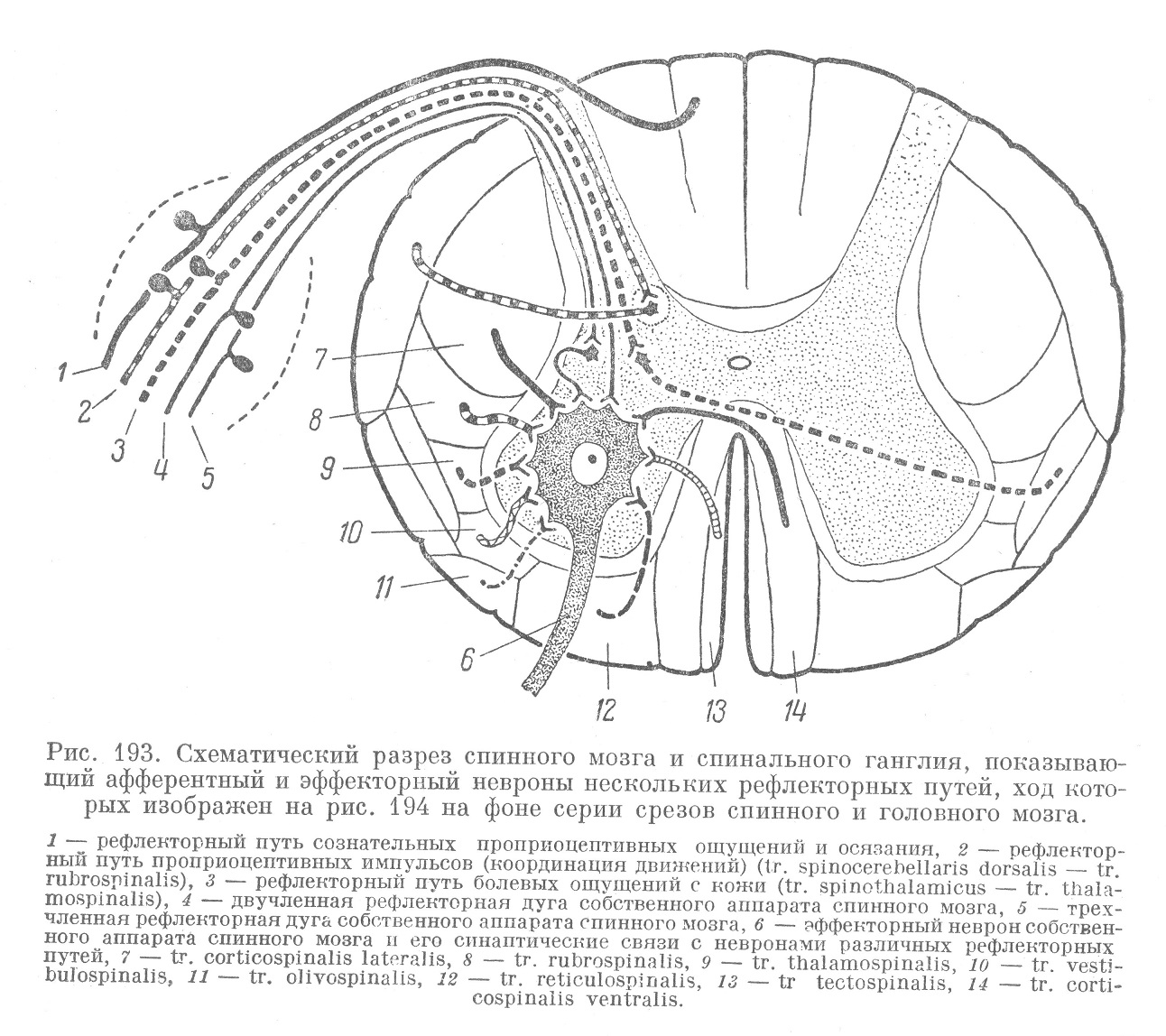
Импульс, идущий по рефлекторному пути, начинающемуся от сетчатки глаза, и передающийся клеткам ядер четверохолмия, направляется далее по нисходящему пути (tr. tectospinalis) в область спинного мозга. Здесь импульс передается двигательным клеткам переднего рога (рис. 193). Этот рефлекторный путь, обозначенный на рис. 193 и 194 поперечно исчерченной мелким штрихом толстой линией, осуществляет защитного характера двигательные акты, возникающие в результате зрительных и отчасти слуховых раздражений.
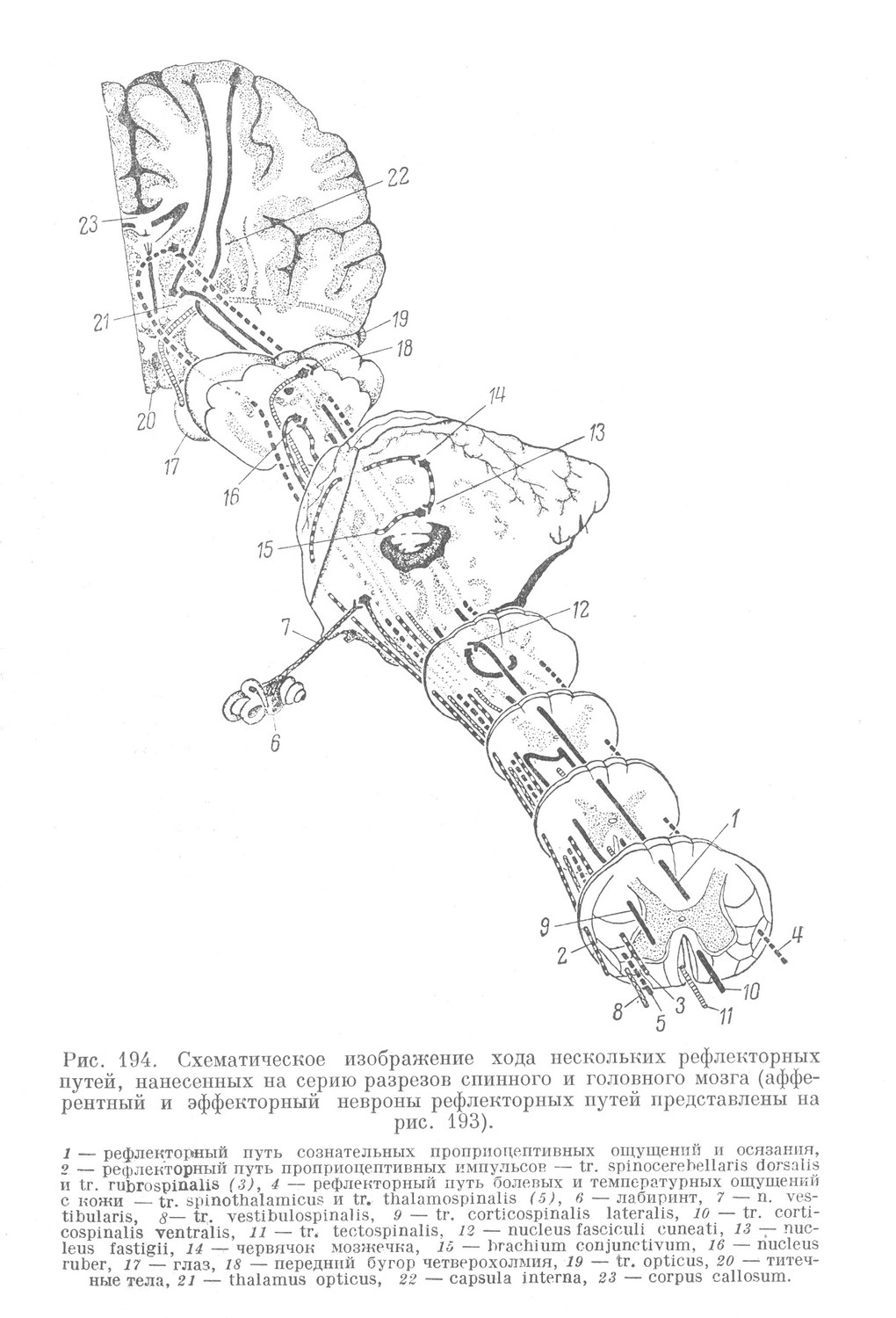
Аппарат связей спинного мозга с корой большого мозга устроен несколько иначе. В белом веществе спинного мозга он представлен восходящими пучками задних столбов и нисходящими (пирамидными) пучками, расположенными в боковых и передних столбах.
Начинается этот путь чувствительными клетками спинальных ганглиев с длинными центральными отростками (невритами). Эти клетки отличаются, по-видимому, более крупными размерами, а в остальном похожи на соответствующие клетки собственного аппарата. Главное отличие их состоит в том, что центральный отросток (на рис. 193, 7, он обозначен толстой линией), войдя через задний корешок, входит в состав заднего столба белого вещества спинного мозга. В некоторых случаях он также дает короткую нисходящую веточку, проникающую в серое вещество и там заканчивающуюся (рис. 192, е). Восходящая ветвь этого отростка имеет значительную длину и тянется в составе задних столбов до продолговатого мозга, где заканчивается в соответствующих ядрах (рис. 194, 12). Из этих ядер импульс переключается на зрительный бугор (рис. 194, 21), а отсюда — на кору большого мозга.
Количество волокон в задних столбах должно увеличиваться по мере приближения к продолговатому мозгу. Наибольшего развития задние столбы белого вещества достигают в шейном отделе, наименее значительны они в хвостовом отделе. Волокна, вступающие в них на более высоких уровнях, все более и более отодвигают к средней септе волокна, идущие из нижних сегментов. В результате в верхних отделах спинного мозга, в заднем столбе различают 2 пучка: один из них, состоящий из более тонких волокон, идущих из нижних отделов, прилегает к задней септе и называется пучком Голля, другой, состоящий из более толстых волокон, идущих из верхних отделов, прилежит к заднему рогу и называется пучком Бурдаха.
Нисходящие связи коркового аппарата начинаются в двигательной области коры больших полушарий и образуются невритами их пирамидных клеток. Эти невриты идут, не переключаясь на другие невроны, непосредственно в передние рога спинного мозга, где и заканчиваются на телах двигательных клеток (рис. 193, 7, 14). Все невриты в своей совокупности образуют 4 пучка, из которых 2 перекрещиваются в продолговатом мозге, а другие 2 остаются на одноименных сторонах. Перекрещенные пучки в спинном мозге проходят в боковых столбах (рис. 192, 14) и называются перекрещенными пирамидными путями (tr. corticospinalis lateralis). Прямые пирамидные пути (tr. corticospinalis ventralis) проходят в передних столбах белого вещества, в их медиальных частях (рис. 192,15). Наиболее значительный объем пирамидные пути имеют в верхних отделах, книзу они постепенно истончаются и теряются в крестцовом отделе спинного мозга.
Особенно сильно развиты пирамидные пути у человека, что находится в соответствии со значительным развитием у него коры большого мозга, откуда эти пути и берут начало.
Таким образом, мы видим, что в основе строения спинного мозга, а вместе с ним и всего цереброспинального отдела нервной системы лежит собственный аппарат спинного мозга, непосредственно связанный с чувствительной и двигательной периферией. Этот процесс находится в двусторонней связи с координирующими центрами головного мозга: во-первых, с низшими и более старыми по происхождению стволовыми центрами различных автоматических функций и, во-вторых, с высшими корковыми центрами сознательных, произвольных движений.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 