Нервная система развивается из нервной трубки и ганглиозной пластинки. Первая в ходе развития дает головной и спинной мозг и отходящие от них черепномозговые и спинномозговые нервы. Из ганглиозной пластинки развиваются спинальные ганглии и связанные с ними проводники, а также аналогичные им ганглии и проводники в области головного мозга и находящиеся за пределами головного и спинного мозга ганглии и проводники вегетативного отдела нервной системы. Из ганглиозной пластинки развиваются также каротидный узел и подобные ему другие образования.
Так как в ганглиозной пластинке и в нервной трубке гистогенетические процессы совершаются более или менее сходно, мы ограничимся изучением процессов, происходящих в нервной трубке.
Стенка нервной трубки в начале своего развития имеет ясно выраженное клеточное строение (рис. 182, а). Благодаря усиленному размножению входящих в ее состав клеточных элементов, имеющих индиферентный эмбриональный характер, стенка нервной трубки постепенно становится многослойной, причем одновременно клеточные границы в ней исчезают (рис. 182, б). Таким образом, на известной стадии развития нервная трубка приобретает строение симпласта (невросимпласта). Вначале все элементы его имеют плотное расположение, в дальнейшем он приобретает губчатый, синцитиальный характер (рис. 182, в).
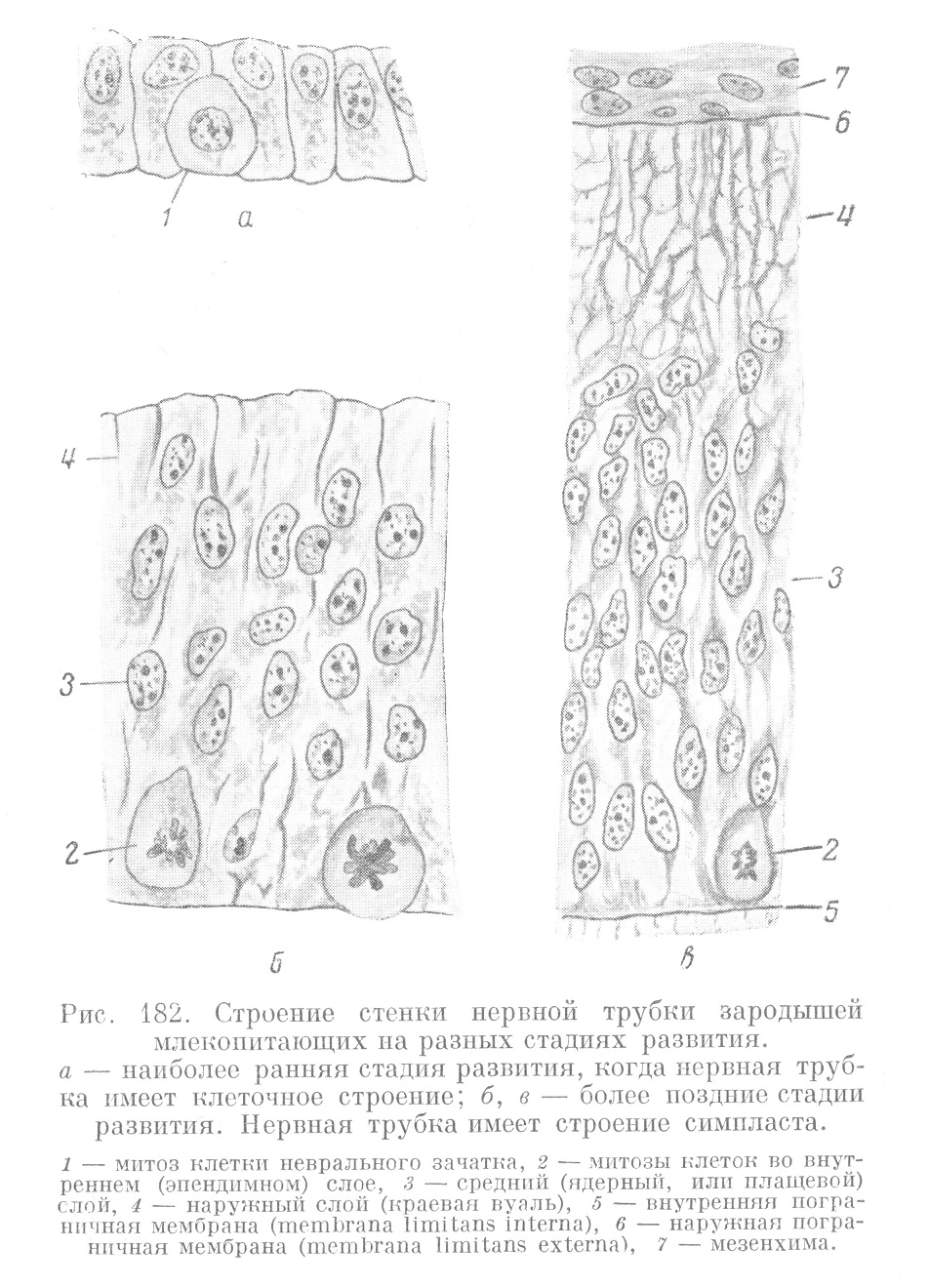
По расположению ядер в синцитиальной стенке различают 3 зоны: внутреннюю эпендимную зону, среднюю, называемую плащевым слоем, и наружную, известную под названием краевой вуали.
В эпендимной зоне, которая сохраняет эпителиальный характер в расположении своих элементов, происходят многочисленные митозы, за счет которых, с одной стороны, увеличивается число ядер в самой эпендимной зоне, а с другой стороны, благодаря смещению ядер вглубь, увеличивается и толща плащевого слоя. Следовательно, в начале развития эпендимный слой выполняет функцию росткового камбиального слоя для всей нервной трубки.
Плащевой слой невросинцития содержит многочисленные ядра, образующиеся в начале развития за счет размножения эпендимных элементов. На более поздних стадиях появляются митозы и в плащевом слое. Краевая вуаль на первых стадиях развития не содержит ядер, они появляются в ней значительно позднее. От окружающей мезенхимы она отделяется ясно выраженной пограничной перепонкой.
Такой невросинцитий является зачатком не только спинного мозга, но и периферических нервов. В дальнейшем в нем путем митозов продолжается размножение индиферентных элементов, все более и более перемещающихся в плащевой слой, и начинается процесс диференцировки, приводящий к образованию в плащевом слое и других участках невросинцития невробластов, из которых позднее развиваются невроны.
Кроме невробластов, в невросинцитии имеются спонгиобласты, из которых развиваются невроглиальные элементы.
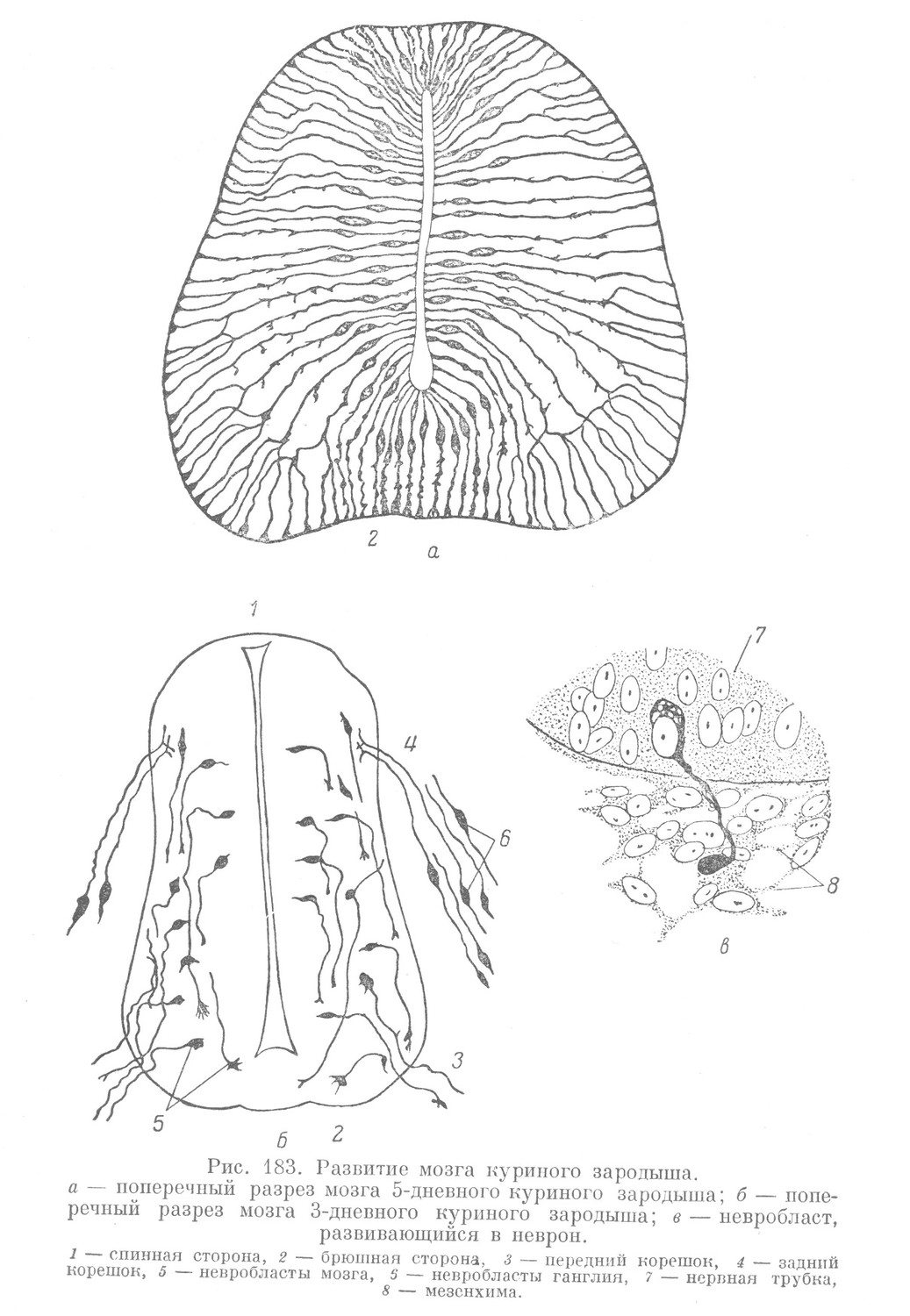
Из спонгиобластов (невроспонгия) образуется основной остов макроглии, носящий вначале эпендимный характер. Элементы этого остова имеют преимущественно радиальное расположение, а содержащие ядра участки лежат вначале в эпендимном слое, позднее по перекладинам синцития они частично смещаются в плащевой слой, а в дальнейшем — в краевую вуаль (рис. 183, а).
Участки невроглиального синцития, остающиеся в эпендимном слое, приобретают форму призматического эпителия и дают начало эпендимной выстилке центрального канала. Вся остальная масса невроспонгия становится макроглией, образующей синцитиальную строму мозга, причем в этой строме в дальнейшем появляются и глиальные фибриллы.
Периферические концы радиальных перекладин глиального синцития обычно имеют пуговчатые утолщения, заканчивающиеся на пограничной перепонке, которая, по всей вероятности, является производной этого синцития. На более поздних стадиях, когда нервные элементы достигнут стадии более или менее высокой диференцировки, из невроглиального синцития выделяются элементы олигодендроглии, которая образует оболочки вокруг тел нервных клеток и нервных волокон белого вещества. В последних миэлин появляется позднее.
Развитие невронов происходит за счет невробластов, которые вначале имеют округлую форму и лишены отростков. На этой стадии развития они еще сохраняют способность к размножению. Последняя утрачивается с того периода, когда они начинают превращаться в невроны, что определяется образованием нервного отростка. Нервный отросток возникает из круглого тела невробласта в форме булавовидного выпячивания и начинает расти в определенном направлении, прокладывая себе путь по невроглиальному синцитию. В таком невробласте очень рано появляются и неврофибриллы, имеющие вначале значительную толщину.
Растущий нервный отросток попадает либо в белое вещество мозга (в краевую вуаль), где он растет по невроглиальному синцитию (из этого синцития в дальнейшем выделяются элементы, дающие начало его миэлиновой оболочке), либо выходит из мозга на периферию. При образовании передних двигательных корешков множество развивающихся невритов в виде пучка выходит из определенного участка спинномозговой трубки и начинает расти по направлению к близлежащему миотому, в котором эти невриты и заканчиваются (рис. 183, б). Вначале пучок невритов не содержит никаких ядер и состоит из голых осевых цилиндров. Позднее между его отдельными волокнами появляются ядра, т. е. клетки, из которых и развиваются в дальнейшем шванновские и миэлиновые оболочки (рис. 183, в).
Вопрос о происхождении этих ядер и клеток (шваняовских клеток) в настоящее время может считаться экспериментально решенным. Шванновские клетки периферических нервов происходят из нервной трубки или из ганглиозной пластинки, т. е. имеют то же происхождение, что невроглия и невробласты, а поэтому и должны быть отнесены к олигодендроглии. Следовательно, никакого принципиального различия между центральными и периферическими волокнами нет. Дендриты у невробластов развиваются сходным путем, но появляются позже невритов.
По такому же типу диференцируются элементы и в ганглиозных пластинках, но здесь невробласты сразу приобретают биполярную форму, так как оба отростка — и центральный и периферический — начинают развиваться почти одновременно. У высших позвоночных биполярные клетки впоследствии преобразуются в псевдоуниполярные.
При развитии нервной системы у высших позвоночных животных все невробласты превращаются в невроны. В нервной системе не остается никаких малодиференцированных элементов, которые могли бы дать в зрелом организме начало новым невронам. Поэтому есть основание предполагать, что основная масса невронов, образовавшаяся в период эмбрионального гистогенеза, сохраняется в нервной системе в течение всей жизни организма. Это положение, основанное на данных изучения гистогенеза и восстановительных способностей элементов нервной системы, ни в какой степени не говорит об ограниченных возможностях к дальнейшему развитию нервной системы высших позвоночных животных, а свидетельствует лишь о высокой ее организации.
Иначе ведут себя элементы глиального синцития, которые сохраняют способность и к размножению и к образованию новых глиальных комплексов, но не обладают способностью превращаться в невробласты.
На основании приведенного описания процессов эмбрионального гистогенеза нервной системы можно уже заранее предвидеть и те регенеративные возможности, которые присущи элементам, входящим в ее состав. Способностью к размножению и к прогрессивному развитию в нервной системе взрослого организма могут обладать, очевидно, только элементы макроглии и микроглии. Нервные элементы, т. е. невроны, целиком восстанавливаться не должны, так как для этого в нервной системе взрослого организма не остается необходимого камбиального источника. Но частичная регенерация в нервных элементах все же происходит. Она заключается в том, что в случае повреждения отростков того или иного неврона последние могут при известных условиях восстанавливаться. Далее, если тело неврона будет приведено в состояние сильного раздражения или выведено из состояния системных отношений, в котором оно находится в норме, характер уже существующих отростков может изменяться и могут появиться новые отростки. Таким образом, содержащая ядро часть неврона, т. е. нервная клетка, сохраняет способность и изменять свою форму, и восстанавливать утраченные части. Поэтому всякое повреждение в центральной нервной системе приводит, во-первых, к регенеративному раздражению глиальных и соединительнотканных элементов, образующих смешанный глиально-соединительнотканный рубец, и, во-вторых — к беспорядочному прорастанию этого рубца регенерирующими нервными отростками поврежденных невронов. Ни восстановления утраченных невронов, ни полного возобновления нарушенных связей при повреждении центральной нервной системы не происходит.
В периферической нервной системе, т. е. в периферических нервах, имеющих только нервные отростки невронов (тела которых лежат в центральной нервной системе или в ганглиях), может произойти и полная регенерация утраченных частей с полным восстановлением их функции.
В экспериментальных условиях регенерацию периферических нервов можно вызвать путем их перерезки. Периферический нерв обычно перерезают на известном расстоянии от мозга. Каждый спинномозговой нерв содержит двигательные и чувствительные волокна, являющиеся отростками двигательных клеток спинного мозга и чувствительных клеток соответствующих ганглиев. Таким образом, нерв после перерезки разделяется на центральный отросток, остающийся в связи с мозгом и ганглиями, и периферический, связь волокон которого с соответствующими нервными клетками оказывается нарушенной.
Непосредственно после операции на месте разреза происходят первичные дегенеративные изменения. Они выражаются в распаде на некотором незначительном расстоянии от места повреждения осевых цилиндров в обоих отрезках — ив центральном и в периферическом. В дальнейшем волокна периферических отрезков, связь которых с телами соответствующих невронов нарушена, претерпевают более глубокие изменения, известные под названием уоллеровского (Waller) перерождения (рис. 184, а, б). Несколько позднее из центральных отрезков начинается регенеративный рост осевых цилиндров, приводящий потом к восстановлению функций периферического отрезка (рис. 184, в).
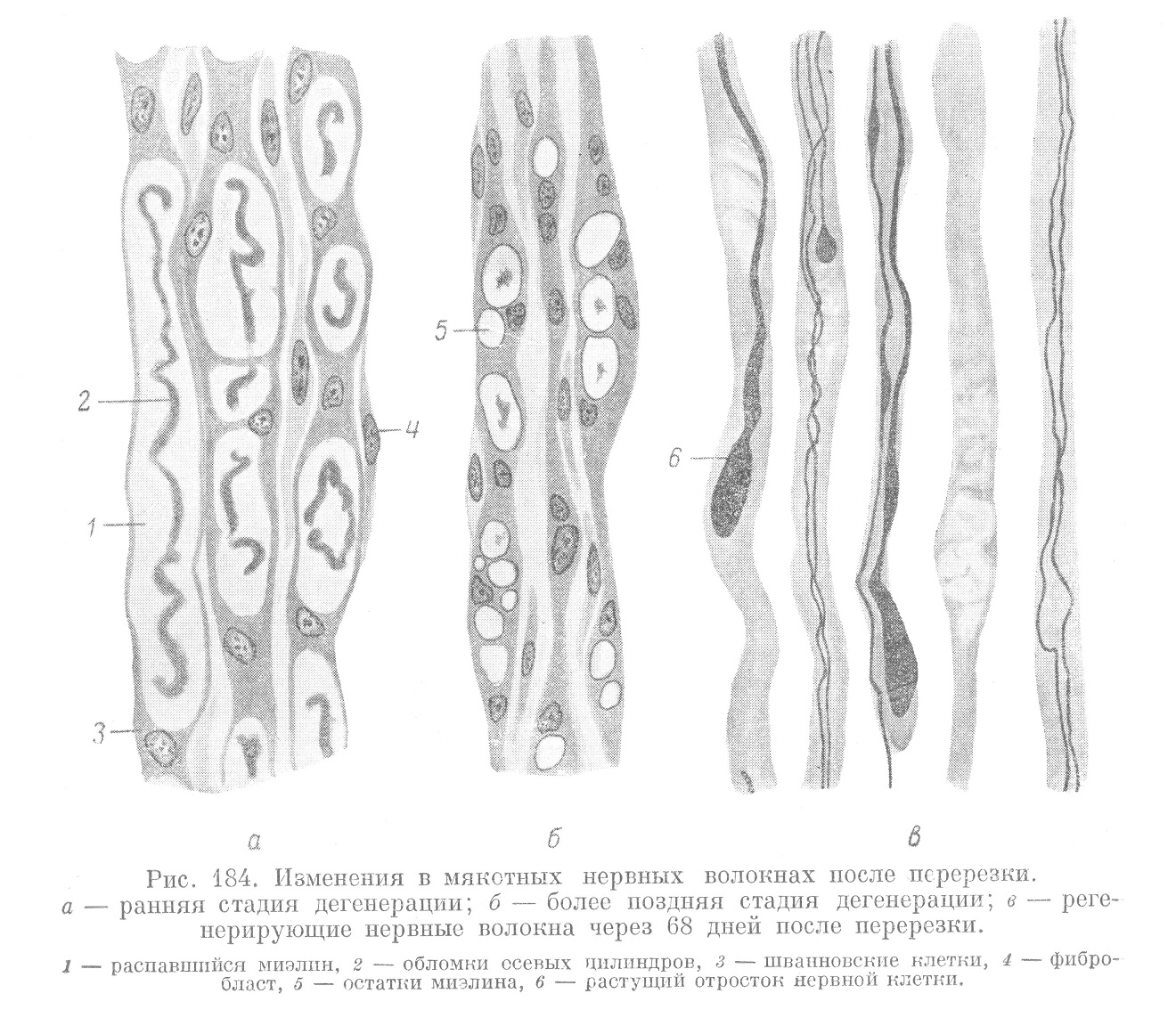
В процессе уоллеровского перерождения осевые цилиндры распадаются на отдельные фрагменты и разрушаются. Одновременно распадается и миэлиновая оболочка, причем миэлин выходит из цитоплазмы шванновских клеток и собирается в крупные капли, в которых оказываются заключенными обломки осевых цилиндров. Цитоплазма шваннов- ского синцития сливается в более плотные массы, куда перемещаются и ядра. Эти ядра располагаются между миэлиновыми каплями, которые остаются окруженными тонким цитоплазматическим ободком. Позднее начинается рассасывание миэлина и обломков осевых цилиндров, в котором принимают участие элементы окружающей соединительной ткани, проникающие через неврилемму внутрь волокна и приобретающие форму макрофагов.
В результате описанных процессов происходит полная резорбция миэлина и распавшихся осевых цилиндров, а волокна периферического отрезка замещаются гомогенными тяжами, образованными цитоплазмой шванновского синцития с ядрами. Такие тяжи одеты неврилеммой. Они получили название лентовидных, или бюнгнеровских, волокон (рис. 184, в). После резорбции миэлина около места перерезки путем митоза начинается усиленное размножение ядер лентовидных волокон и разрастание шванновского синцития. Разрастающиеся шванновские синцитии выходят в рубцовую ткань, образующуюся на месте разреза, и начинают расти в виде переплетающихся тяжей в направлении центрального отрезка. Таким образом, дефект между центральным и периферическим отрезками выполняется тяжами глиальной ткани, происходящей из шванновских элементов периферического отрезка.
Одновременно возникают регенеративные разрастания на концах осевых цилиндров волокон центрального отрезка. Эти концы булавовидно расширяются, в них образуется разрыхленная сеть неврофибрилл, и они начинают расти в сторону периферического отрезка, сопровождаемые элементами шванновского синцития. При этом дело не ограничивается лишь разрастанием только концевого отдела осевого цилиндра. На протяжении известного участка от осевого цилиндра отходят многочисленные боковые веточки, которые также имеют булавовидные утолщения и начинают расти, загибаясь обычно в сторону места перерезки. Таким путем концевые отделы осевых цилиндров центральных отрезков преобразуются в пучки усиленно растущих и разветвляющихся волоконец. Последние врастают в глиальный рубец, образовавшийся на месте перерезки и субстанционально связанный с бюнгнеровскими волокнами периферического отрезка. В результате часть веточек попадает в волокна периферического отрезка и начинает расти в их шванновском синцитии уже по старому руслу, после чего дальнейшее образование боковых веточек прекращается; разветвления, не нашедшие себе правильного пути, подвергаются обратному развитию, а попавшая в лентовидное волокно веточка растет до места нервного окончания, где возникает соответствующий концевой аппарат.
Нервные веточки, врастающие в лентовидные волокна периферического отрезка, превращаются в их осевые цилиндры; в шванновском синцитии появляется миэлин, и, в конце концов, восстанавливается нормальная структура мякотного волокна. Одновременно восстанавливается и функция соответствующих периферических органов.
Рассмотренный пример регенерации периферических нервов ясно указывает и на значение невроглии, и на значение окружающей соединительной ткани, и на регенеративные возможности нервных элементов. Вместе с тем этот пример, равно как и эмбриональный гистогенез периферических невронов, является весьма веским аргументом в пользу невронной теории.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 