Кора больших полушарий на всем своем протяжении имеет неодинаковое строение, что обусловливается различным функциональным значением разных ее отделов. Рассматривая кору как совокупность анализаторов (точнее, их центральных частей — ядер), И. П. Павлов определил расположение последних как динамическую структуру, в которой резкие границы между анализаторами отсутствуют и зоны соседних анализаторов заходят друг за друга. Между ядрами анализаторов имеются участки коры, принадлежащие не одному, а различным анализаторам. Они осуществляют лишь простейшие, элементарные анализы и синтезы, тогда как в ядрах анализаторов совершаются высшие анализы и синтезы.
Для удобства изучения строения и расположения невронов в коре последнюю принято условно делить на несколько слоев. В наиболее развитых отделах коры больших полушарий (например, в передней центральной извилине) (рис. 226, а) различают 7 слоев:
1) молекулярный слой,
2) слой малых пирамид,
3) слой средних пирамид,
4) наружный слой больших пирамид,
5) слой звездчатых клеток,
6) внутренний слой больших пирамид,
7) слой полиморфных клеток.
На некоторых участках коры отдельные из перечисленных слоев развиты слабо или вообще отсутствуют.
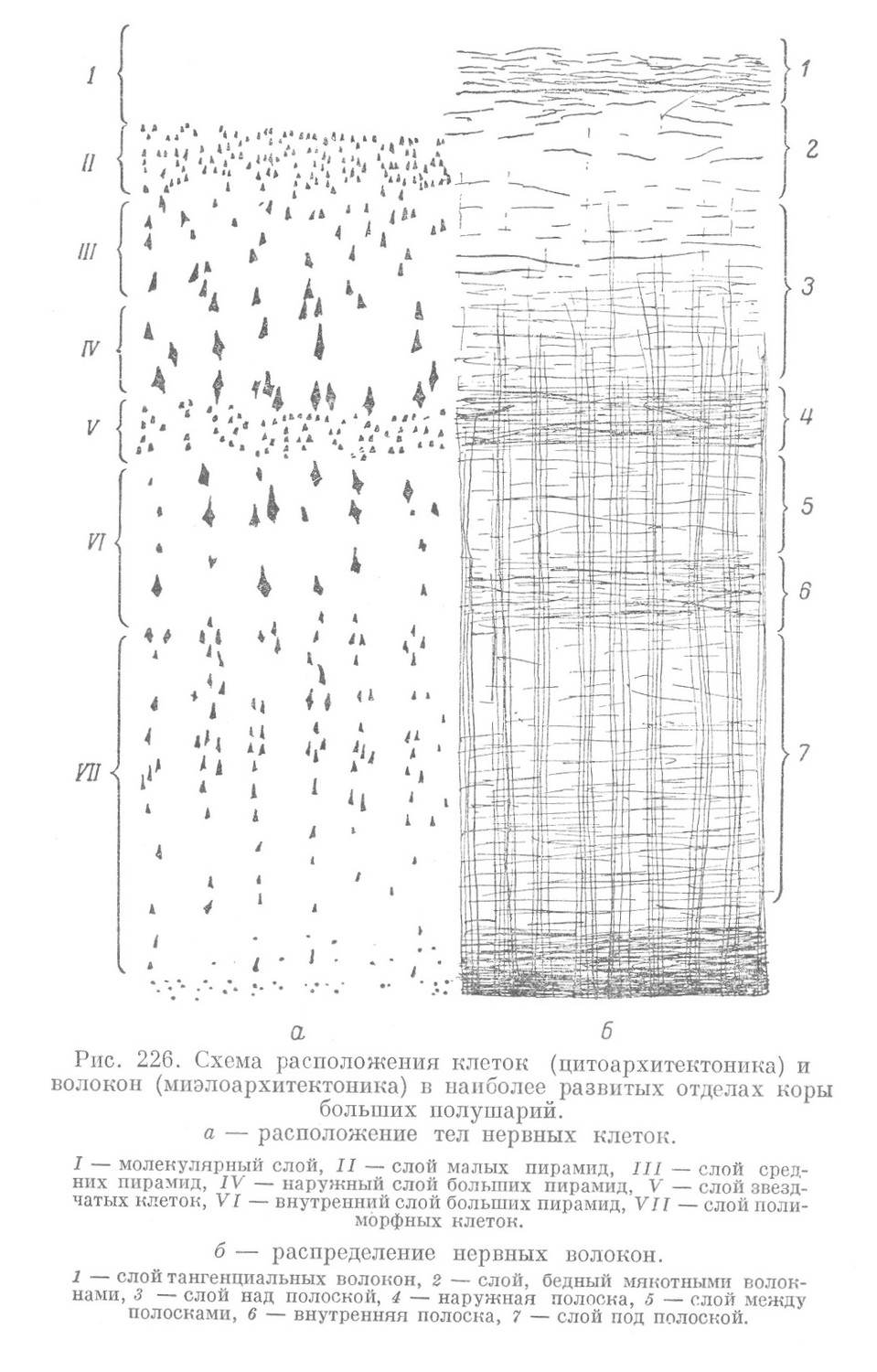
Миелиновые волокна проходят в горизонтальном и вертикальном направлениях и количество их в разных местах коры различное (рис. 226, б). Поэтому при изучении хода миэлиновых волокон принято различать в коре 7 слоев:
1) слой тангенциальных волокон,
2) слой, бедный мякотными волокнами,
3) слой над полоской,
4) наружная полоска,
5) слой между полосками,
6) внутренняя полоска,
7) слой под полоской.
Границы перечисленных слоев с указанными выше слоями невронов не совпадают.
Строение коры можно хорошо представить себе лишь в том случае, если расположение клеток (цитоархитектоника) и расположение волокон (миелоархитектоника) рассматриваются одновременно.
В функциональном отношении в коре следует различать следующие образования:
1) центростремительные (афферентные) волокна, входящие в кору из белого вещества,
2) эфферентные невроны, аксон которых выходит из состава коры в белое вещество,
3) промежуточные (ассоциативные) невроны, образующие сочетательные системы коры.
Центростремительные волокна принято делить на проекционные и ассоциативные. Первые (рис. 227), идущие от зрительного бугра и коленчатого тела, являются центростремительными проводниками различных анализаторов. Они несут в кору нервные импульсы как из внешней, так и из внутренней среды организма. В области внутренней полоски они дихотомически делятся,образуя сплетение;волокна от последнего идут к поверхности коры, и на уровне наружной полоски снова делятся, образуя другое афферентное сплетение. От него отходят концевые веточки, теряющиеся в слое над полоской.
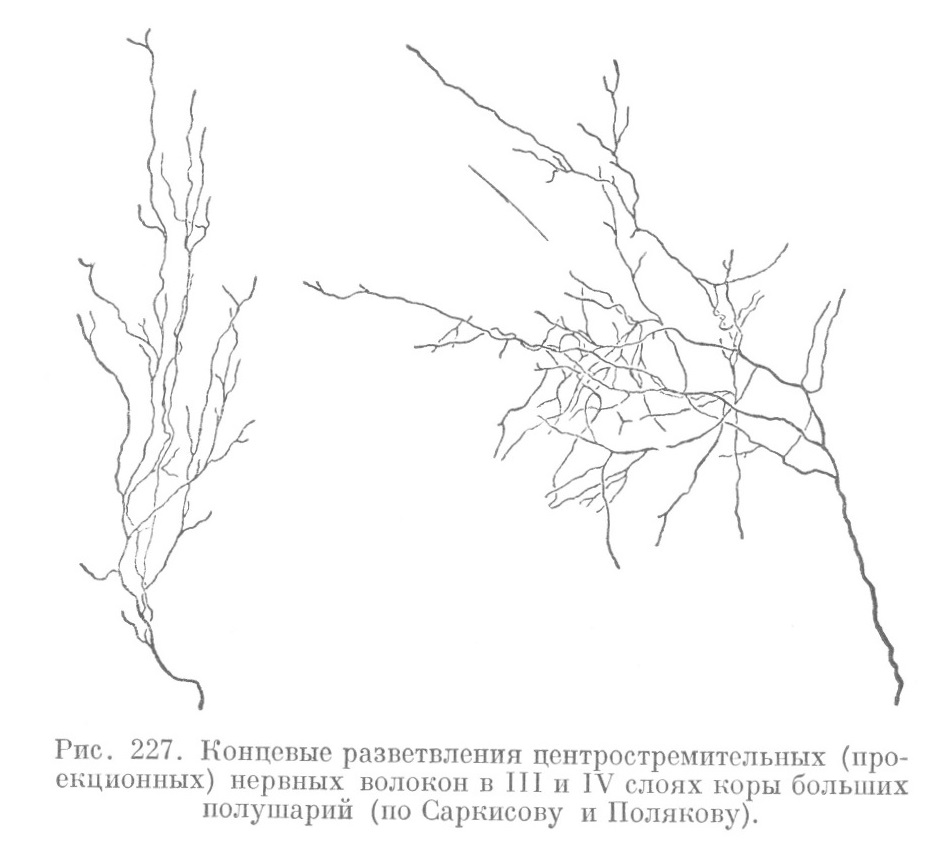
Ассоциативные волокна бывают короткие и длинные. Первые соединяют кору соседних участков полушария, тогда как вторые соединяют отдаленные участки коры одного полушария или участки, расположенные в разных полушариях. Основная масса коротких ассоциативных волокон заканчивается в глубоких слоях коры. Длинные волокна распространяются и в другие слои коры до третьего слоя включительно.
Функцию эфферентных невронов выполняют пирамидные и веретенообразные клетки. Они имеют различные размеры и делятся на 2 группы. Одни из них располагаются во втором, третьем и четвертом слоях и образуют своими аксонами указанные выше ассоциативные пути.
Другие, расположенные в пятом, шестом и седьмом слоях, своими аксонахми образуют центробежные проводники, по которым нервные импульсы от коры направляются к ядерным центрам низшего отдела нервной системы.
К промежуточным, или вставочным, невронам относятся звездчатые, паукообразные, кустовидные и горизонтальные клетки.
Их аксоны не выходят за пределы серого вещества. Первые 3 вида обнаруживаются в различном количестве во всех слоях коры, за исключением молекулярного. В последнем лежат горизонтальные клетки. В коре больших полушарий человека, по сравнению с животными, количество промежуточных клеток особенно велико.
Имеются указания, что из 14 млрд, невронов, насчитываемых в коре человека, указанные выше клетки составляют больше половины. Они образуют различные сочетательные системы, расположенные как в горизонтальном направлении между невронами в пределах одного слоя,так и в вертикальном между невронами различных слоев коры. Имеются сочетательные системы и более сложного порядка, объединяющие деятельность невронов, расположенных в обоих указанных направлениях. Промежуточные невроны в системе коры, по-видимому, имеют исключительное значение в распространении импульса как в первой фазе, когда возбуждение иррадиирует и распространяется по всей коре, так и во второй фазе, когда оно концентрируется в определенном пункте коры. В последнем случае оно передается эффекторным невронам, посылающим импульсы к соответствующим центрам низшего отдела нервной системы и в двигательные ядра спинного мозга, что в результате и проявляется в форме определенной деятельности организма.
Межневрональные отношения в коре, ввиду их большой сложности, до последнего времени изучены недостаточно.
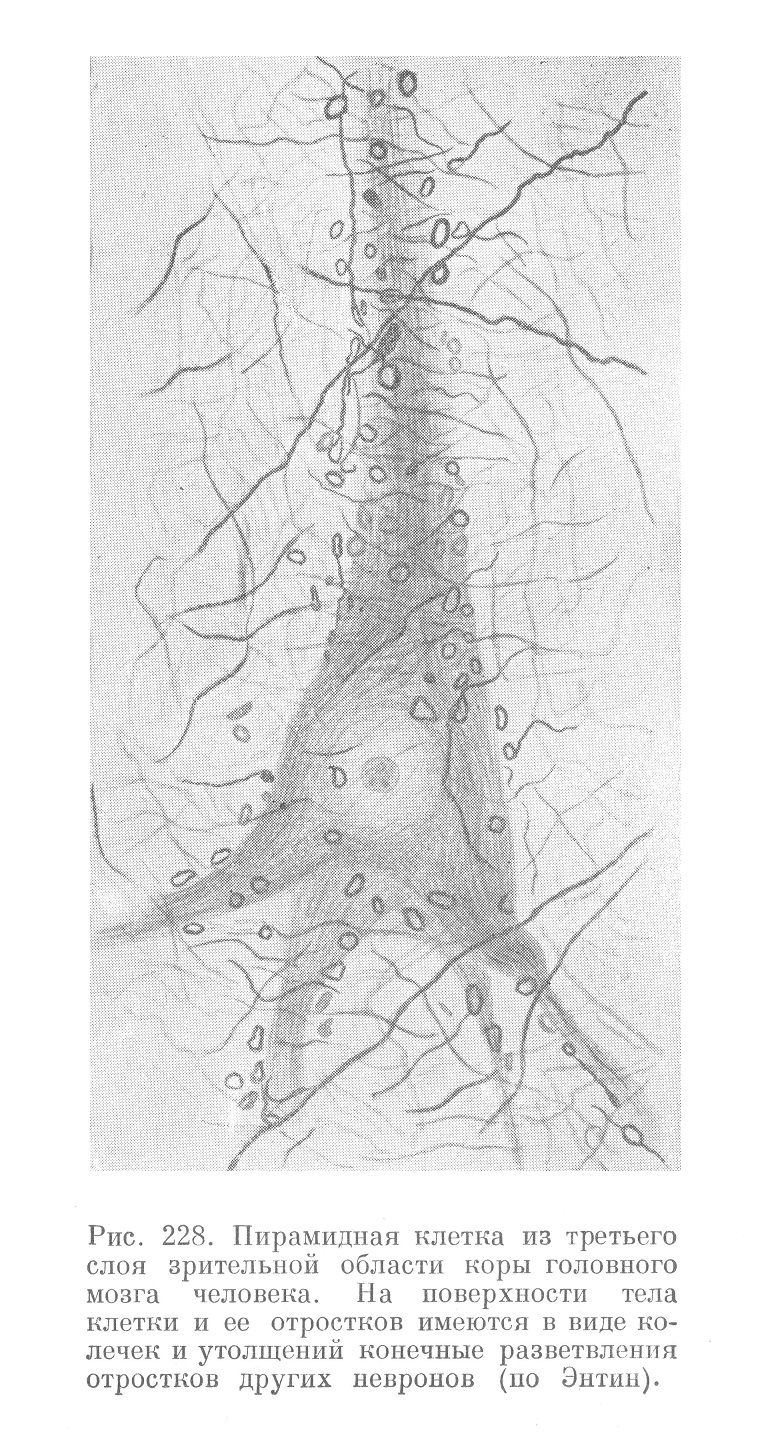
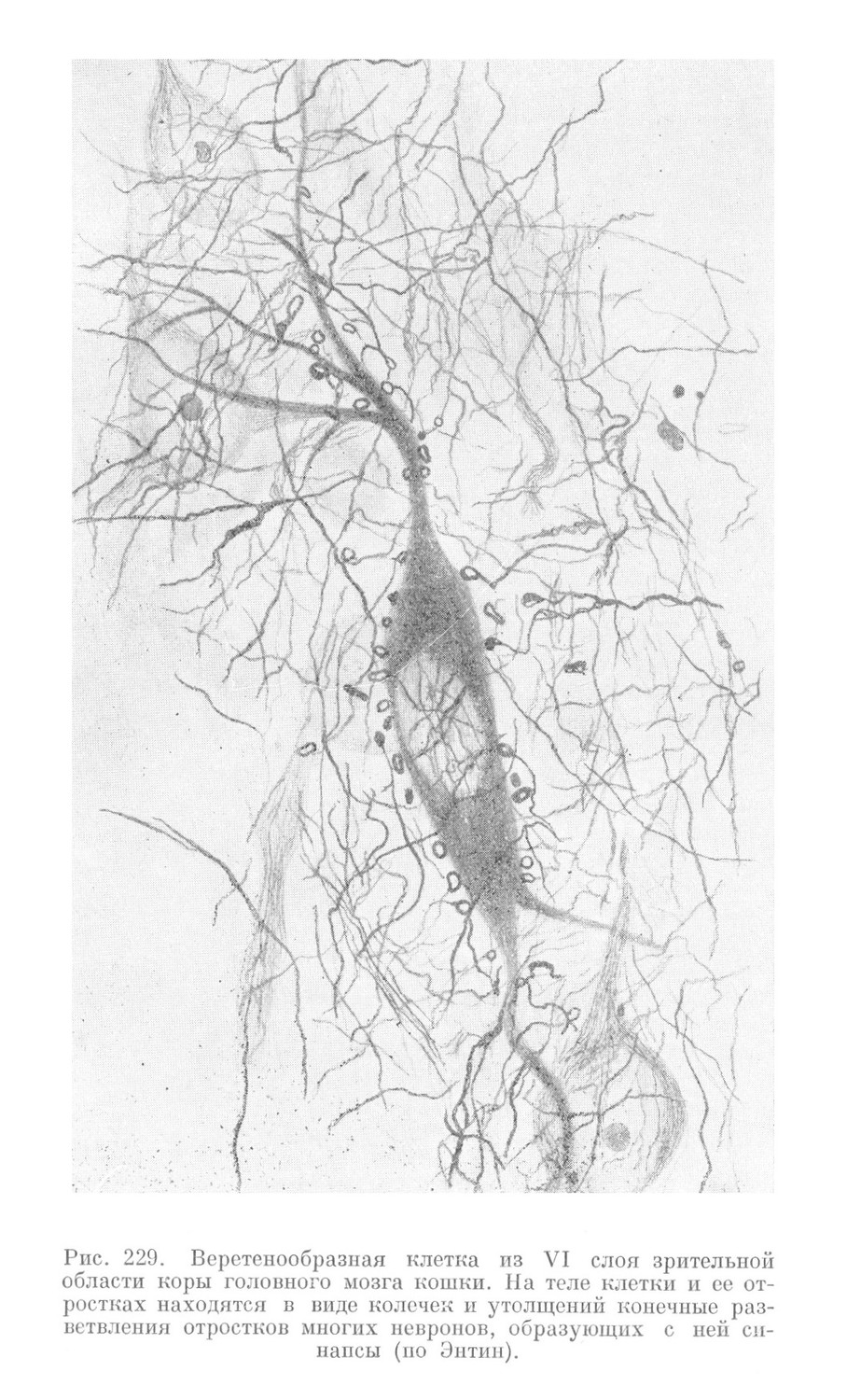
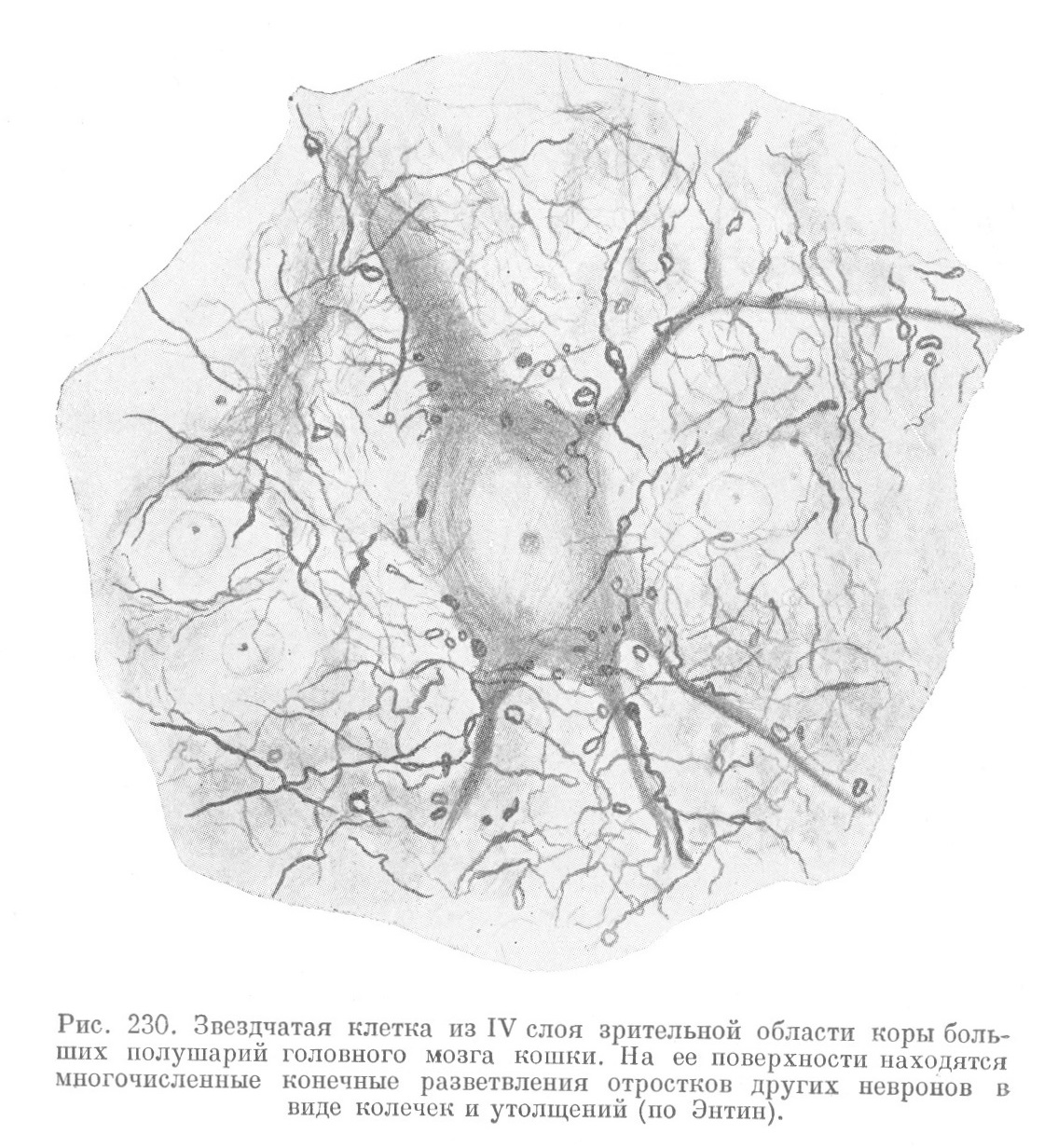
Места контактов между невронами — синапсы — имеют простое или і более сложное строение. В первом случае разветвление аксона одного неврона завершается утолщениями в виде пуговок или петель на поверхности тела и дендритов другого неврона (рис. 228, 229 и 230). Во втором случае разветвления аксона образуют вокруг тела другого неврона нежную сеть. На месте контакта аксона с дендритами описаны также мелкие шипики, которые отходят от дендрита на значительном протяжении и у разных невронов выражены различно.
Со стороны мягкой мозговой оболочки в состав коры проникает большое количество мелких артерий мышечного типа. Большая часть их разветвляется и переходит в густую сеть капилляров, расположенных в поверхностной или глубокой части коры. Другие артерии достигают белого вещества мозга и на различной глубине его переходят в более рыхлую сеть капилляров. Кровь из капиллярной сети коры и белого вещества собирается в вены, которые, сопровождая артерии, выходят из мозга в мягкую мозговую оболочку.
Густота капиллярной сети в разных слоях коры неодинакова. Она значительно варьирует в пределах даже одного и того же слоя. Это связано, по-видимому, с неодинаковым расположением и различной функцией невронов. Особое внимание обращает на себя взаимоотношение между капиллярами и крупными невронами (рис. 231).
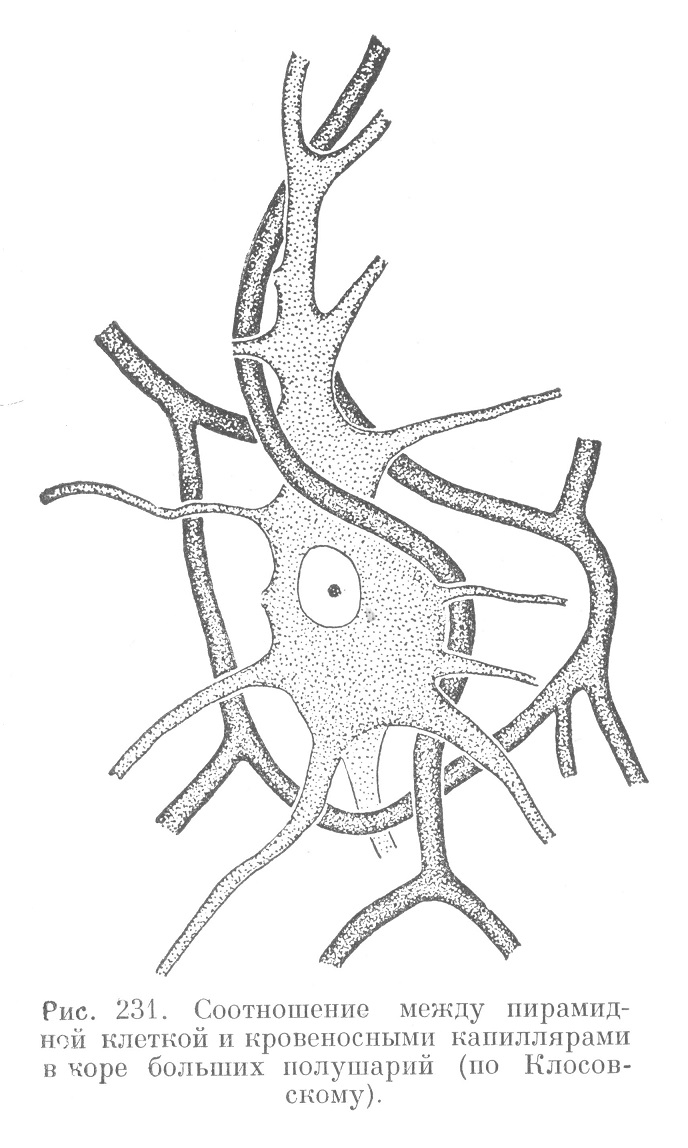
Вокруг последних капилляры образуют петли, составляющие в целом сеть, которая в отдельных местах тесно прилегает к неврону. Здесь тело неврона отделено от стенки капилляра лишь тонким слоем цитоплазмы клеток олигодендроглии, выполняющей как трофическую, так и разграничительную функции.
Вместе с сосудами из мягкой мозговой оболочки в кору больших полушарий проникает большое количество нервных стволиков. Последние представляют продолжение нервного сплетения, лежащего в мягкой мозговой оболочке. Эти стволики идут спирально вокруг сосудов и, разветвляясь, образуют в их адвентиции сплетение. Отдельные нервные волокна сплетения завершаются в мышечной оболочке сосудов свободными окончаниями. Они являются эффекторными (вазомоторными) проводниками, несущими импульсы к сосудистой мускулатуре. Описаны также эфферентные (чувствительные) проводники, выходящие из состава мозга и лежащие, так же как и двигательные проводники, по ходу сосудов. Они завершаются свободных древовидно разветвленных рецепторами, имеющими вид окончаний (рис. 232).
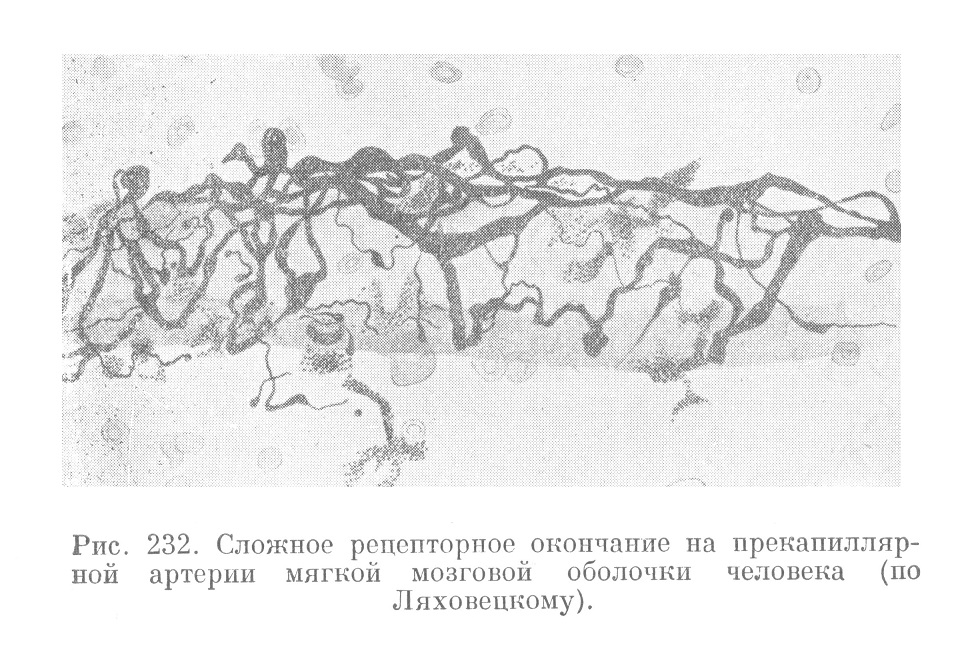
Все описанные нервные проводники и нервные окончания коры, расположенные по ходу ее кровеносных сосудов, относятся к вегетативному отделу нервной системы. Они связаны с очень тонкой и сложной функцией регуляции кровообращения в высшем отделе нервной системы.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 