Деление клеток следует рассматривать как одно из важных проявлений жизнедеятельности ткани. Оно постоянно сопровождает развитие всех тканей. Но так как ткани развиваются в течение всей жизни организма, то и деление клеток наблюдается на всех этапах онтогенеза. Однако интенсивность размножения клеток разных тканей на протяжении онтогенеза изменяется. Наиболее бурно происходит размножение клеток в эмбриогенезе, при образовании эмбриональных зачатков и последующем развитии из них тканей. В постэмбриональном периоде в пограничных тканях и тканях внутренней среды размножение клеток идет достаточно интенсивно и связано с постоянным обновлением структуры указанных тканей.
В соматической мускулатуре и в нервной системе деление основных тканевых элементов — поперечно-полосатых мышечных волокон и нервных клеток — составляет исключение. Оно наблюдается лишь в первой половине гистогенеза. Мышечные волокна соматической мускулатуры и нервные клетки пребывают в организме весьма длительное время. Основная масса их функционирует в течение всей его жизни. Развитие их выражается не столько в изменении морфологического строения (что по сравнению с пограничными тканями и тканями внутренней среды выражено сравнительно слабо), сколько в постоянном обновлении и развитии их химического состава. В тесной связи с этим идет развитие их функции.
Из сказанного следует, что интенсивность размножения клеток стоит в прямой связи с продолжительностью их жизни и вместе с тем зависит от характера функции, которую выполняет тканевая система.
Значительная продолжительность жизни нервных клеток у высокоорганизованных животных и у человека связана с длительной эволюцией, с усложнением организма и его нервной системы. Если вопреки действительности предположить, что у названных представителей в нервной системе в течение всей жизни организма клеточный состав обновляется, т. е. постоянно происходит гибель невронов и на их месте возникают новые невроны, то в этом случае непременно наблюдалась бы непрерывно протекающая перестройка и изменение нервных связей.
Если учесть, что восстановление нервных волокон после их повреждения происходит весьма медленно (в течение нескольких месяцев, а иногда и лет), то в предполагаемом случае постоянно нарушалась бы интегрирующая роль нервной системы в разных областях тела.
Мышечные волокна соматической мускулатуры являются эффекторным аппаратом нервной системы. Способности к размножению у них ограничены. Однако, по сравнению с нервной системой, в соматической мускулатуре имеются более значительные восстановительные способности, о чем подробнее будет сказано при рассмотрении строения мышечной системы.
Указанные выше биологические свойства приобретены элементами нервной и мышечной системы у высших животных и человека в ходе длительной эволюции. У низкоорганизованных животных обновление структуры нервной системы и мускулатуры происходит в широких пределах. В качестве примера можно привести восстановление целостности организма у дождевого червя из сравнительно небольшого участка тела. Здесь происходит образование новых невронов и мускулатуры из малодиференцированных элементов.
Различают два способа размножения клеток: митоз (кариокинез), или непрямое деление, и амитоз, или прямое деление. Последнее многие исследователи до недавнего времени неправильно трактовали не как размножение клеток, а как одно из проявлений жизнедеятельности клеток, находящихся в абортивном состоянии и потерявших в значительной степени свою жизнеспособность.
Митоз или кариокинез
Митоз впервые был описан в растительных клетках И. Д. Чистяковым в 1874 г. В животных клетках его первым исследовал П. Н. Перемежко в 1879 г. В дальнейшем эта форма деления клеток была подвергнута всестороннему исследованию.
Клетка, вступившая в процесс деления пребывает в ткани в морфологической, функциональной и генетической связи с соседними элементами. В начале митоза происходит некоторое обособление клетки. Она теряет значительную часть межклеточных мостиков, исчезает синцитиальная связь с соседними элементами и с промежуточным веществом. Вследствие нарушения указанных связей изменяется и форма клетки; ее цитоплазма как бы собирается в комочек. Диференцированные части цитоплазмы, например реснички на поверхности некоторых эпителиальных клеток, тонофибриллы и другие образования, пропадают. Включения в цитоплазме также исчезают. Вязкость цитоплазмы изменяется, она становится более плотной.
Перечисленные изменения клетки перед митозом свидетельствуют о том, что она переходит в период пониженного жизненного обмена, в недеятельное состояние. В таком случае период жизни клетки между митозами, так называемый интеркинетический период, следует считать рабочим состоянием, когда обменные реакции и функции клетки протекают более бурно.
Процесс митоза для удобства принято условно разделять на 4 фазы: профазу, метафазу, анафазу и телофазу. Все 4 периода переходят друг в друга постепенно и в целом составляют единый процесс (рис. 28 и 29).
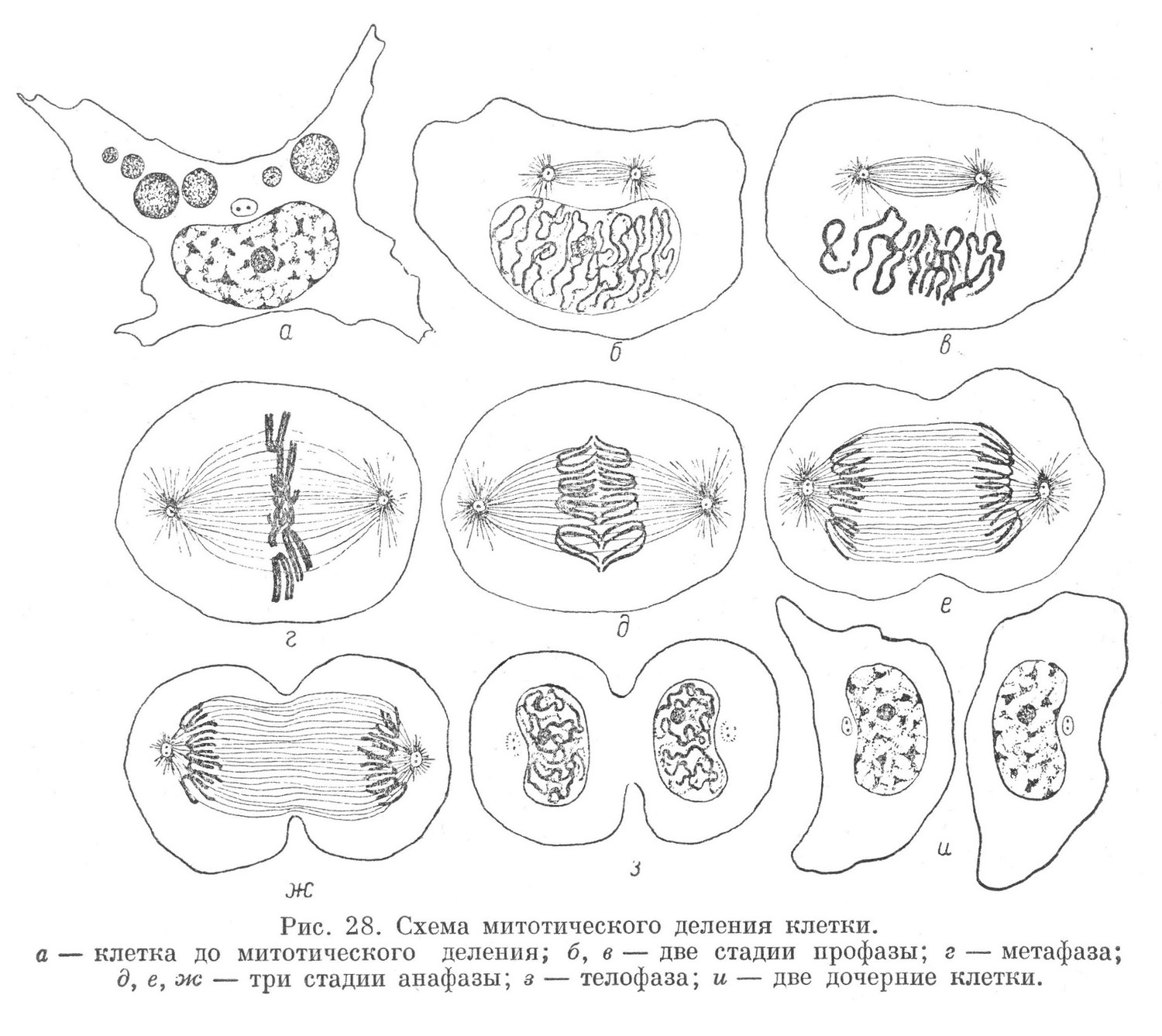
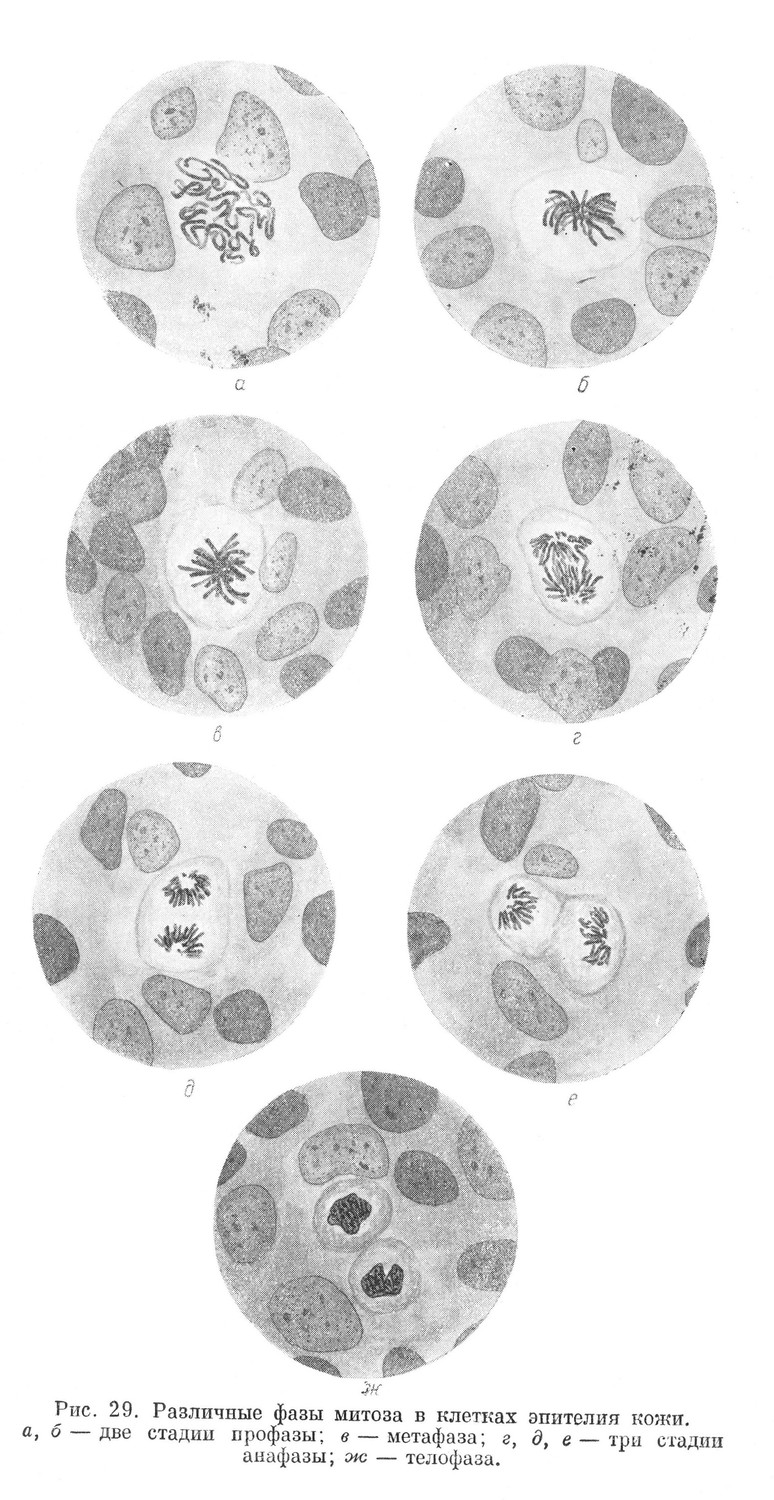
В профазе наблюдаются изменения структуры ядра. Обычная ядерная сеть, отмечаемая на гистологических препаратах, исчезает. В митотичиски делящейся клетке на месте ядерной сети образуется структура, имеющая вид плотного клубка, образованного тонкой хроматиновой нитью, хорошо окрашивающейся ядерными красками.
Позднее эта нить утолщается, и клубок становится более рыхлым. Эту стадию называют рыхлым клубком. Вслед за этим исчезают ядрышко и оболочка ядра, содержимое которого без особой грани соединяется с цитоплазмой. Нить, образующая клубок, ясно разделяется на отдельности, имеющие преимущественно вид петель, называемых хромозомами (рис. 28,в). Одновременно с изменением ядра происходит превращение клеточного центра. На стадии плотного клубка центриоли расходятся по направлению к полюсу ядра. Центродесмоза и прилегающий к ней участок цитоплазмы образуют тонкие нити, так называемое ахроматиновое веретено. На стадии рыхлого клубка центриоли еще более удаляются друг от друга, вместе с этим удлиняется ахроматиновое веретено.
После полного исчезновения ядерной оболочки хромозомы оказываются вначале свободно лежащими в цитоплазме и расположенными беспорядочно. В дальнейшем, в метафазе, хромозомы располагаются более закономерно в плоскости, проходящей через экватор, при этом они устанавливают связь с нитями ахроматинового веретена. Все хромозомы располагаются таким образом, что изогнутые части их обращены к продольной оси клетки, а концы — к периферии. Если наблюдать хромозомы с полюса, то они оказываются расположенными в виде звезды. Этот момент митоза называют поэтому стадией материнской звезды (рис. 28,г и 29,6).
В дальнейшем происходит продольное разделение каждой хромозомы на две части, которые в течение некоторого времени остаются вместе.
Расхождение их происходит в начале анафазы. Половинки хромозом расходятся к полюсам. При расхождении их некоторую роль играет, по-видимому, укорочение связанных с ними ахроматиновых нитей веретена. Вначале расходятся изогнутые части половинок хромозом, позднее — их концы. У каждого полюса образуется скопление в определенном порядке расположенных хромозом, которые при рассматривании с полюса напоминают звезду. Эту стадию митоза обозначают как стадию дочерних звезд (рис. 28,е, ж и 29, д, ё). Затем наступает последний период митоза — телофаза.
В телофазе строение дочерних звезд нарушается, хрохмозомы располагаются в беспорядке, образуя рыхлый клубок. Затем выявляется оболочка ядра и ядрышко. Рыхлый клубок становится плотным. Центриоли разделяются каждая на две. Ахроматиновое веретено пропадает. Разделение тела клетки начинается еще в анафазе, в результате появления циркулярной борозды (рис. 28, с, ж). В телофазе эта борозда постепенно углубляется и материнская клетка окончательно делится на две новые дочерние клетки.
Хондриом и внутренний сетчатый аппарат во время митоза сохраняются в клетке без значительных изменений. Лишь в некоторых случаях сетчатый аппарат расчленяется на отдельности. При разделении тела клетки указанные органоиды механически делятся между дочерними клетками.
За телофазой следует период, когда в клетке происходит реконструкция ядра. Строение его становится обычным для неделящейся клетки, находящейся в периоде интеркинеза. Вначале тела клеток представляются еще более или менее обособленными от окружающих элементов ткани. Позднее, в ходе развития, они постепенно приобретают свойственную им форму, при этом устанавливают морфологические связи как с соседними клетками, так и с промежуточным веществом. В клетках появляются свойственные им специализированные структуры, в цитоплазме накапливаются различные включения. Клетка вступает в деятельность (рабочий период).
Изменение клетки во время митоза достаточно отчетливо выступает при жизни. Процесс деления можно заснять микрокинематографически. При просматривании кадров фильма, показывающих митоз, ясно видно возникновение хромозом, их расщепление и различное расположение в разные периоды деления клетки, а также их исчезновение в дочерних клетках. Они бесструктурны и состоят из однородной гомогенной массы. Достаточно отчетливо выступает веретено, которое, как показывают экспериментальные исследования, при жизни клеток обладает большей плотностью по сравнению с окружающей цитоплазмой. Однако нити веретена при жизни не выявляются. Отчетливо выступает разделение тела клетки, дальнейшее изменение формы дочерних клеток и установление их- связи с соседними тканевыми элементами.
Продолжительность отдельных фаз митоза различна. Митотическое деление мезенхимных клеток в тканевых культурах протекает во времени следующим образом. Профаза длится от 30 до 60 минут, метафаза — 2— 10 минут, анафаза — 2—3 минуты, телофаза — 3—12 минут; период, в течение которого завершается реконструкция ядра клетки, длится от 30 до 120 минут. Наиболее продолжительными являются начальный и конечный периоды митоза, т. е. то время, когда возникают хрѳмозомы и когда они исчезают при переходе клетки в интеркинетическое состояние.
Скорость течения митоза обусловливается влиянием внешней среды и различным функциональным состоянием ткани. Причинами, замедляющими или ускоряющими митоз, являются температура, действие лучистой энергии и др.
Большое влияние на митотическое деление клеток оказывает нервная система. Механизм этого действия следует понимать, по-видимому, не как непосредственную передачу нервного импульса митотически делящейся клетке через нервный проводник, а как влияние, оказываемое нервной системой на ткань в целом, вследствие чего в ней изменяется метаболизм, а это в свою очередь сказывается как на количестве возникающих митозов, так и на скорости их течения.
Многополюсные митозы
Сравнительно редко в тканях встречаются митозы, имеющие необычный вид. В них отмечается не два, а три и более полюсов (рис. 30). Такие митозы называются многополюсными. В отдельных случаях они наблюдаются в костном мозге, при делении гигантских клеток, называемых мега- кариоцитами. Встречаются они в эпителиальных тканях, культивируемых вне организма. Половая клетка морского ежа, оплодотворенная в экспериментальных условиях несколькими сперматозоидами, также делится путем многополюсного митоза. Особенно часто многополюсные митозы встречаются в делящихся клетках эпителиальных опухолей.
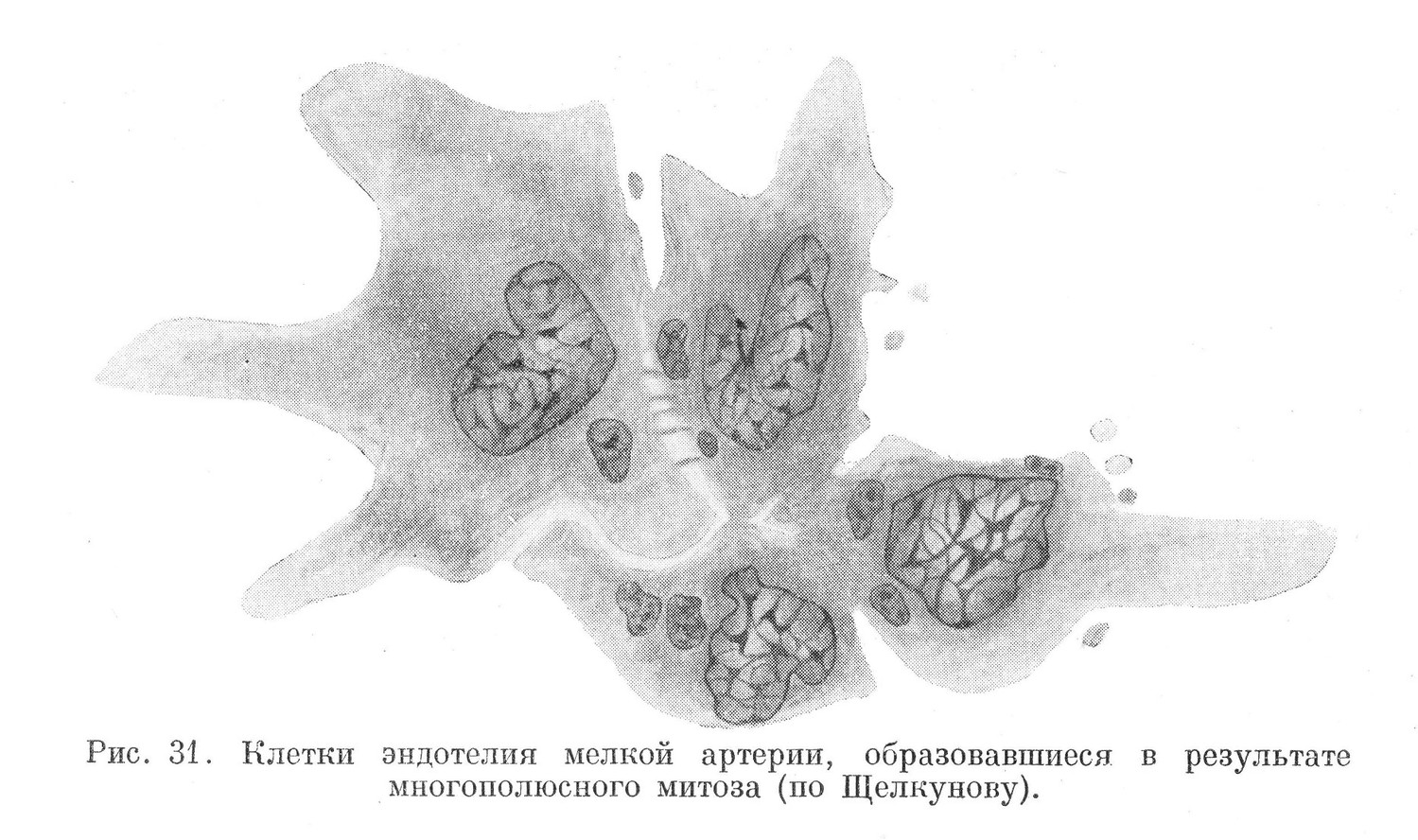
Протекает многополюсный митоз так же, как и обычный двухполюсный. В нем различают те же фазы. Особенностью его является то, что две и более центриоли, имеющиеся в делящейся клетке, расходятся в разные стороны, и каждая из них вместе с цитоплазмой образует полюс. Между полюсами возникает сложное ахроматиновое веретено. В метафазе хромозомы располагаются во многих плоскостях. Все ойи по отношению к различным полюсам будут экваториальными. Вследствие этого фигура материнской звезды принимает сложное очертание. В анафазе половинки расщепившихся хромозом в различном количестве расходятся к полюсам. В дальнейшем у каждого полюса формируется ядро, размеры и форма которого бывают различны.
Разделение тела клетки происходит не всегда. В том случае, когда оно не происходит, многополюсный митоз приводит к образованию многоядерной гигантской клетки. Но если разделение совершается, то возникают сразу 3—4 и более клеток (рис. 31). Число последних может быть и меньшим, чем количество возникших ядер. Это бывает тогда, когда часть дочерних клеток содержит по 2 и более ядер.
Появление многополюсного митоза прежде всего связано с наличием в клетке многих центриолей. Они возникают или путем деления ранее существующих в клетке центриолей, или вносятся другими клетками, например при оплодотворении женской половой клетки несколькими сперматозоидами. Причины, побуждающие центриоли к разделению, остаются невыясненными. Основной причиной является, повидимому, изменение внешних условий, в которых пребывает ткань.
Прямое деление, или амитоз
Кроме митотического деления, клетки могут размножаться и путем амитоза, или прямого деления. Это явление в жизнедеятельности клеток было известно задолго до митоза. Первоначально его рассматривали как единственную форму размножения клеток. G открытием митоза и на основе последующего более глубокого изучения этого процесса амитозу, как форме размножения клеток, стали придавать все меньше и меньше значения. Более того, амитоз стали рассматривать как неполноценное деление клетки или как одно из проявлений жизнедеятельности клетки, потерявшей в значительной степени свою жизнеспособность. К этому выводу исследователей побуждало, с одной стороны, то, что амитоз часто встречается в тех клетках ткани, которые достаточно высокодиференцированы, а с другой стороны, простота самого процесса деления, несложность изменений амитотически делящейся клетки, дающей в результате перешнуровывания нередко две различные по величине клетки, а иногда и одну многоядерную клетку. В связи с этим амитоз рассматривали лишь как явление роста ядра, связанное с функциональным состоянием клетки. Подобного мнения придерживались также те исследователи, которые разделяли теорию Вейсмана — Моргана, утверждавшую, что передача наследственных признаков обусловлена генами, заложенными в хромозомах. Так как хромозомы во время амитоза не выявляются и разделение наследственного вещества (генов) при амитозе не происходит, то и самый амитоз, по их мнению, нельзя рассматривать как размножение клеток.
Амитоз встречается во всех четырех типах тканей. В их жизнедеятельности он занимает различное место и в морфологическом отношении не является строго однородным. Одна разновидность амитоза состоит в разделении всей клетки и образовании двух, а иногда и большего количества дочерних клеток. Другая — ограничивается разделением лишь ядра материнской клетки на два и более новых ядер. Цитоплазма при этом не разделяется, вследствие чего образуется дву- и многоядерная клетка (рис. 32). Если разделение ядер повторяется, то возникает симпласт.
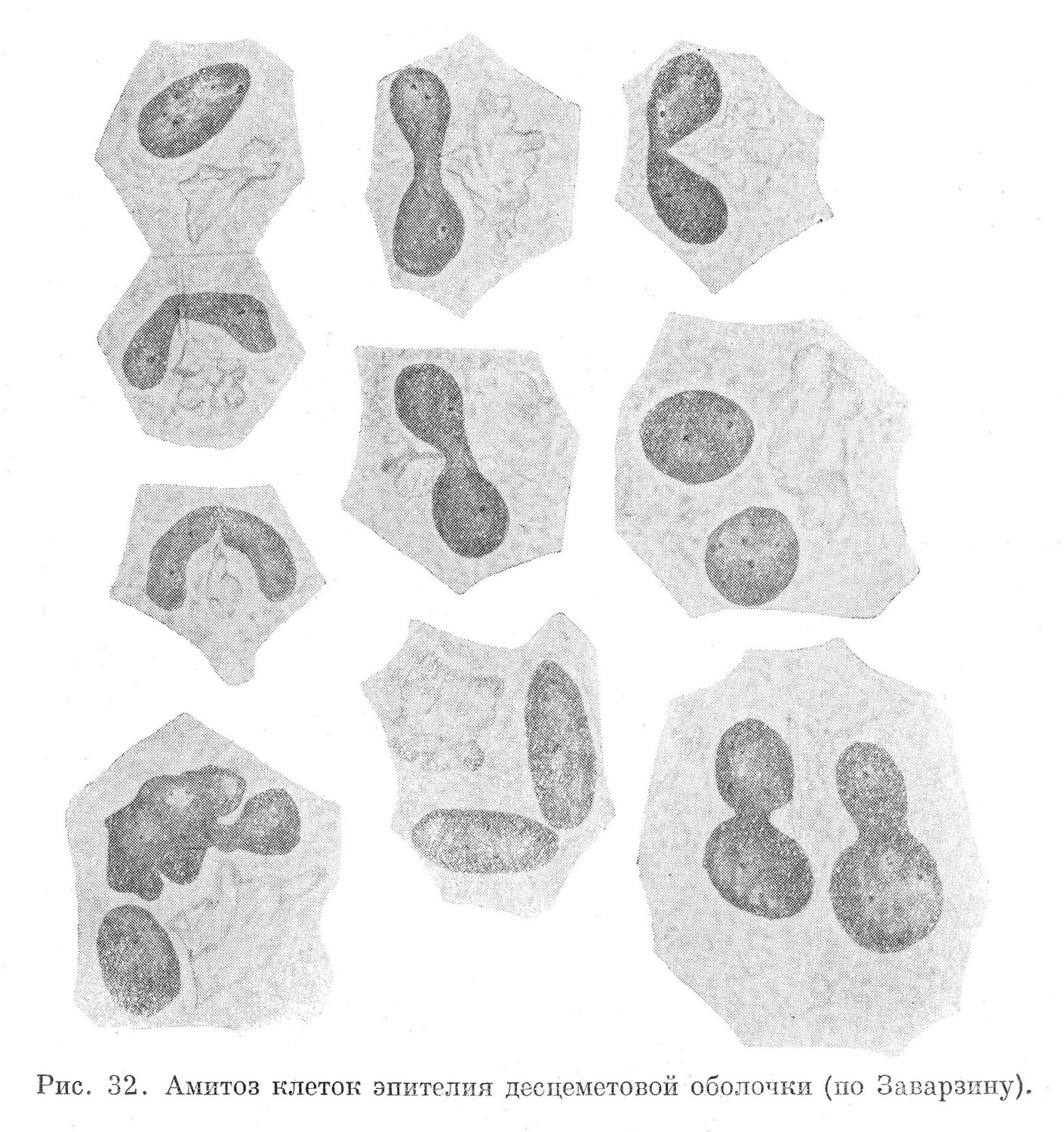
В отличие от митоза клетка, вступающая в амитоз, не проходит подготовительной стадии, выражающейся в ее обособлении в составе ткани и отделении как от соседних клеток, так и промежуточного вещества. Включения из цитоплазмы не во всех случаях исчезают. Внутриклеточные метаплазматические образования остаются без видимых изменений, а иногда могут развиваться (рис. 33).
Наиболее часто встречающаяся форма амитоза, приводящая к образованию двух и более дочерних клеток, начинается с того, что на поверхности ядра появляется циркулярная борозда, которая, постепенно углубляясь, разделяет его на два новых ядра (рис. 32).
Структура ядра ни в период, предшествующий амитозу, ни во время амитоза заметных изменений не претерпевает. В некоторых случаях перед амитозом наблюдается расхождение ядрышек к краям по продольной оси ядра, при этом, если в ядре имелось одно ядрышко, то оно предварительно разделяется. По сравнению с ядром разделение тела клетки запаздывает. Оно начинается или тогда, когда перешнуровывание ядра зашло достаточно далеко, или даже после полного разделения ядра. Цитоплазма распределяется между дочерними клетками то более, то менее равномерно. Образуются две новые клетки.
Как отклонение от описанного и наиболее часто встречающегося амитоза отмечается такое деление клетки, когда в результате перешнуровывания ядра одновременно образуется более двух новых ядер. Последующее разделение цитоплазмы совершается между двумя, а иногда и тремя дочерними клетками.
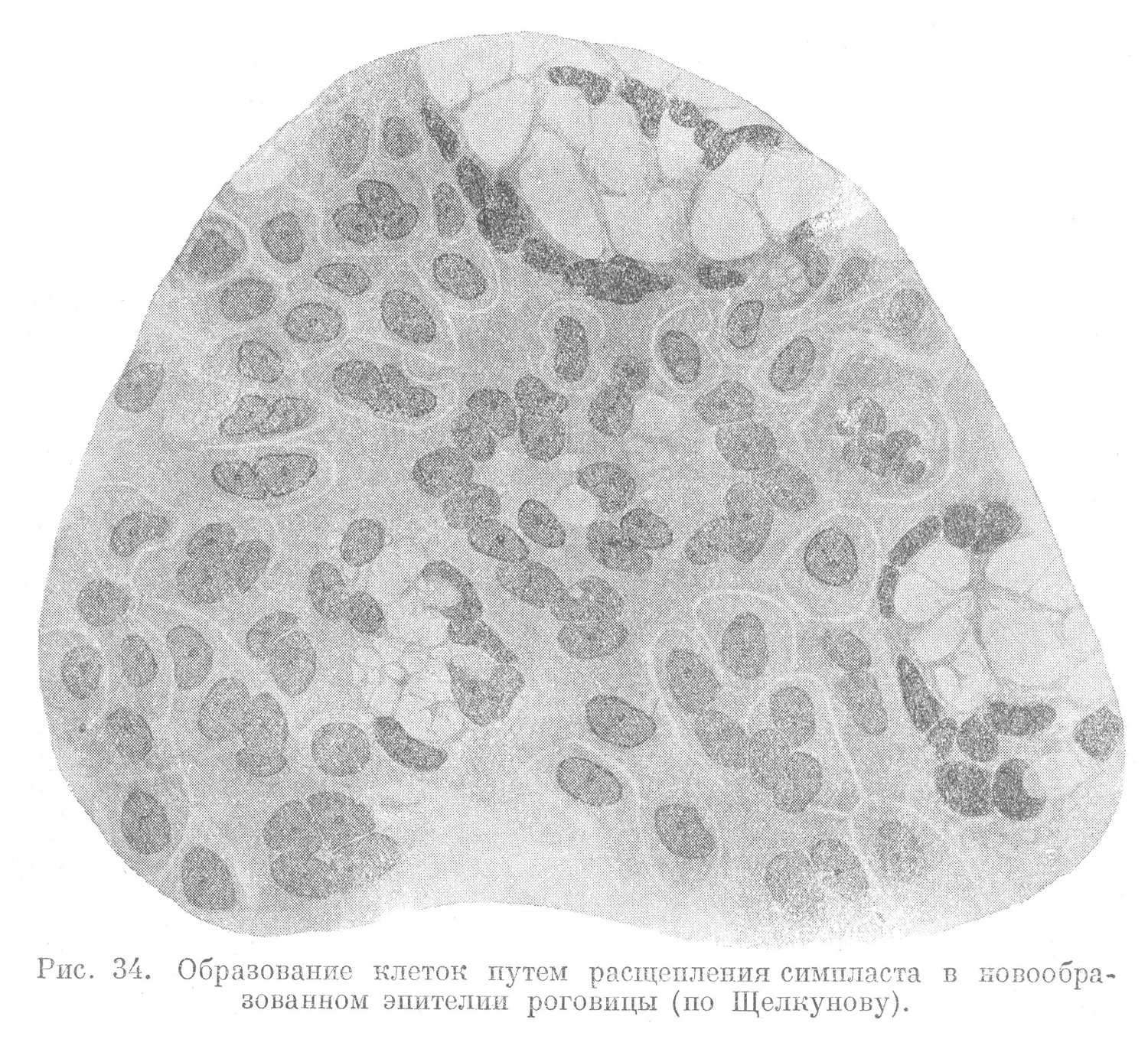
Другая форма амитоза, приводящая к образованию одной многоядерной клетки, сопровождается разделением лишь ядра. Ядро в таких случаях дает два или несколько новых ядер, чаще неодинаковой формы и различных размеров. Наряду с крупными получаются и совсем мелкие ядра, вследствие чего разделение ядра материнской клетки в таких случаях нередко имеет характер неправильного почкования. Возникшая таким образом многоядерная клетка имеет значительные размеры и в некоторых случаях достигает гигантской величины (рис. 33). Иногда перешнуровывание ядер может неоднократно повторяться. В результате этого, вместе с увеличением количества ядер, увеличивается и масса цитоплазмы. В возникшем симпласте без особого порядка разбросано значительное количество ядер. Иногда наблюдается расчленение симпластов на отдельные клеточные элементы, имеющие одно или несколько ядер. При этом между клеточными территориями симпласта (участками, образованными ядрами и окружающей их цитоплазмой) выявляются узкие бледно окрашивающиеся щели. На гистологических препаратах эти щели остаются неокрашенными.
Они придают вычленяющимся из симпласта клеткам различные очертания. Такие клетки могут быть одноядерными и многоядерными (рис. 34).
В других случаях симпласты сохраняются более продолжительное время и могут обнаруживать дальнейшую диференцировку, в результате которой в них появляются внутриклеточные метаплазматические образования. Такие явления можно наблюдать при развитии поперечнополосатых мышечных волокон скелетной мускулатуры.
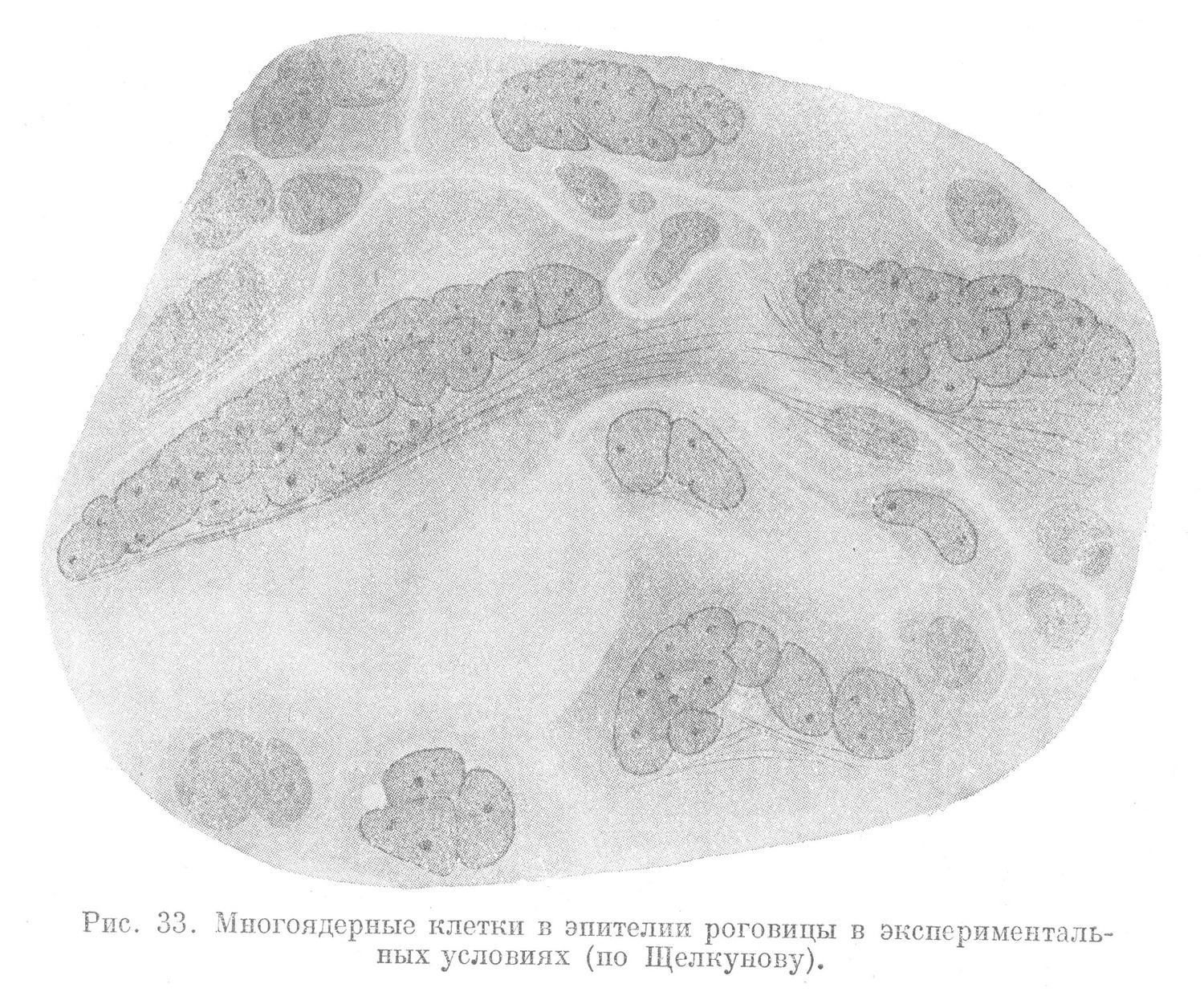
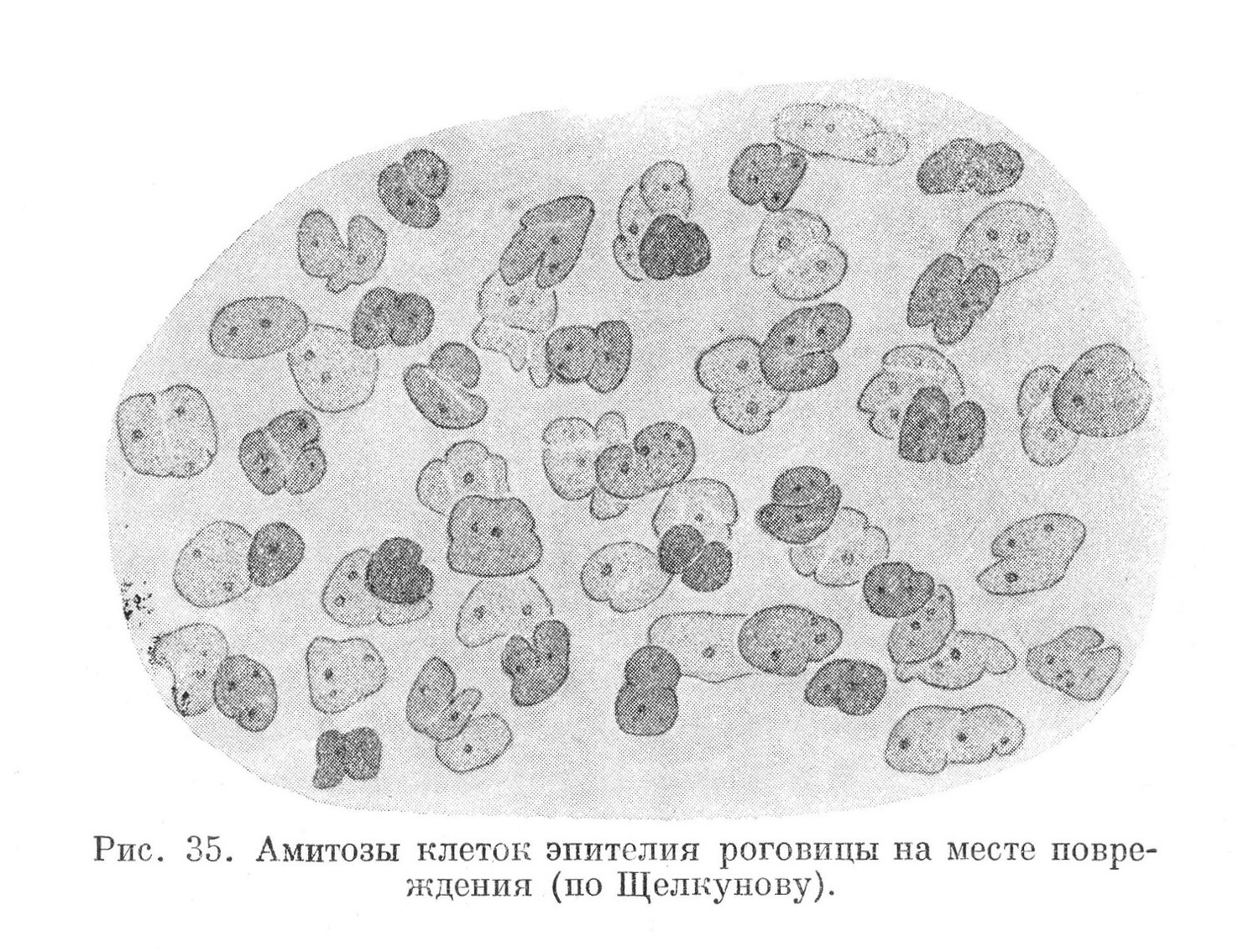
Амитотическое деление свойственно клетке на всех этапах ее развития. В одних тканях при обычных условиях эта форма деления встречается в достаточно высокоразвитых клетках, в других — амитотически делятся и молодые, малодиференцированные клетки. В пограничных тканях, имеющих прямой контакт с внешней средой (например в кожном эпителии), амитозы в обычных условиях встречаются в клетках более диференцированных и отсутствуют среди молодых, малодиференцированных. Однако, если последние подвергнуть непосредственному влиянию внешних факторов (путем удаления поверхностных слоев эпителия), то они делятся исключительно амитотически (рис. 35). Для эпителия вторичной полости тела, пребывающего в условиях внутренней среды организма и не имеющего прямого контакта с внешней средой, основной формой размножения клеток является амитоз.
При восстановлении гладкой мускулатуры на месте повреждения развитие новой мышечной ткани сопровождается преимущественно амитозами. То же наблюдается и при восстановительных процессах соматической мускулатуры; в таких случаях амитозы занимают исключительное место, а митозы встречаются лишь в единичных случаях. Таким образом, амитоз в жизни различных тканей занимает неодинаковое место, и одного подхода к оценке этого явления в жизни клеток в неоднотипных тканях быть не может. Для правильного понимания этого процесса в каждом отдельном случае необходимо исходить из функционального состояния ткани и условий, в которых она пребывает.
Учитывая, что амитозы в различных тканях встречаются как в молодых, так и в высокодиференцированных клетках и что они появляются в тканях и при раздражении, некоторые исследователи предложили разделить все амитозы на 3 группы: генеративные, реактивные и дегенеративные. Однако подобное разделение было бы формальным, так как генеративные амитозы, наблюдающиеся в молодых клетках при росте и развитии тканей, одновременно могут быть и реактивными. Наоборот, реактивные амитозы могут одновременно быть и дегенеративными, иногда предшествуя дегенерации клетки.
Что касается отношения амитоза к митозу в онтогенезе клетки, то этот вопрос остается еще недостаточно изученным. Одни исследователи полагают, что обе формы размножения могут чередоваться, другие — считают начальной формой размножения клеток митоз, а последующей — амитоз. Ни один из этих взглядов до последнего времени не получил достаточного научного обоснования.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 