Органоидами называют постоянные, высокодиференцированные образования цитоплазмы, связанные, главным образом, с осуществлением клетками их общих функций, например внутриклеточного обмена веществ, секреторной и других видов деятельности. К органоидам относятся: хондриом, внутренний сетчатый аппарат, клеточный центр.
Хондриом
В конце прошлого и в начале настоящего столетия в цитоплазме клеток различных тканей многими исследователями была обнаружена структура, имеющая зернистый или фибриллярный характер. Ее назвали хондриомом. В одних клетках хондриом состоит из мелких отдельно лежащих зерен хондриозом, в других — из нитей — хондриомитов. Иногда отдельности хондриома имеют вид зернистых нитей, называемых митохондриями (рис. 15).
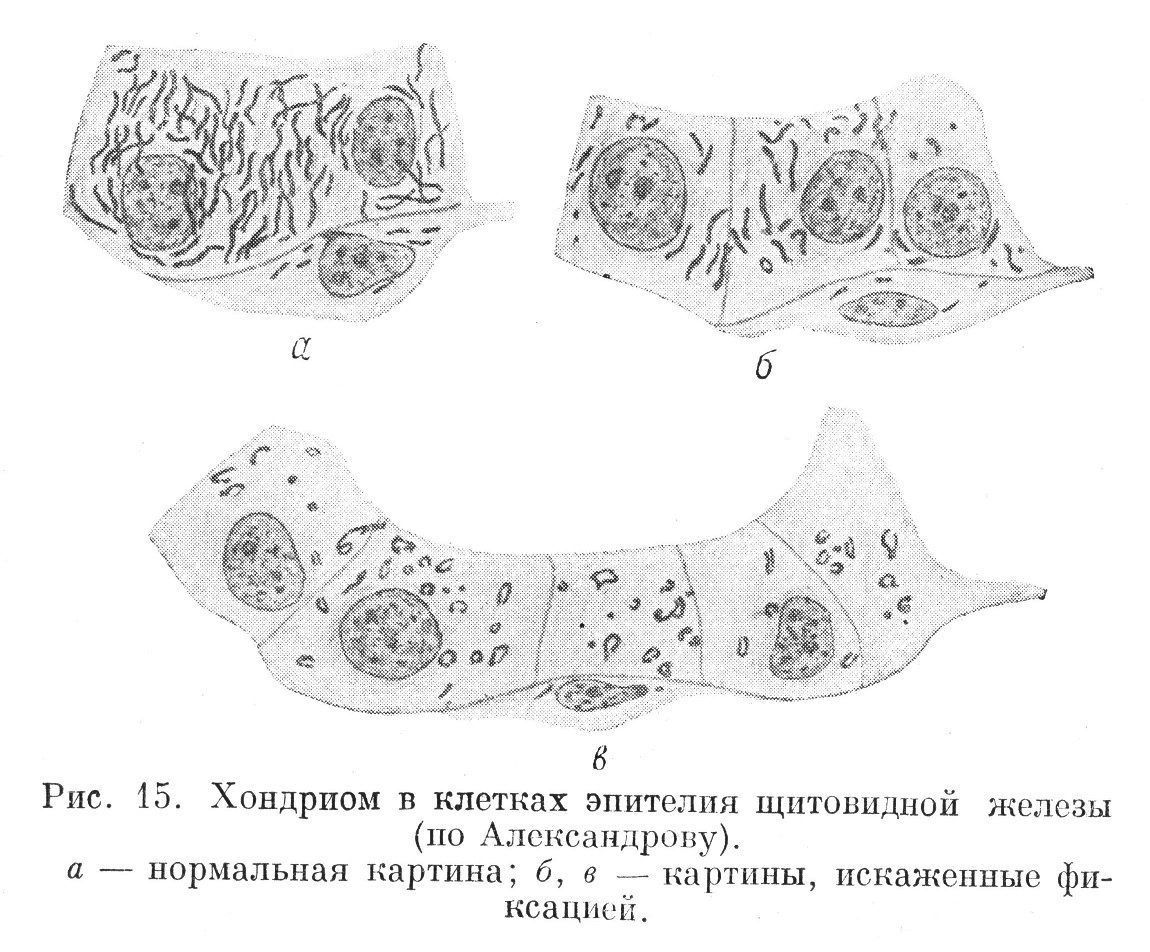
При некоторых условиях хондриом можно наблюдать в цитоплазме живых клеток (как животных, так и растительных). В животных клетках при жизни хондриом окрашивается краской «янусгрюн». Под влиянием внешних условий хондриом может быстро изменяться. Его отдельности утрачивают типичную форму, и хондриом частично или полностью исчезает. При обработке ткани фиксирующей жидкостью, содержащей кислоту или спирт, хондриом разрушается. Химический состав хондриома, по-видимому, сложный, в него входят липоиды и белки. Соотношение между этими компонентами изменяется в зависимости от условий, в которых пребывает клетка, и от ее функции. В некоторых клетках при известном их состоянии в хондриоме преобладают белки. В таких случаях он сохраняется также и после обработки ткани кислыми фиксаторами.
В клетках различных тканей хондриом выражен неодинаково. Более того, даже в одной и той же ткани, но в разных клетках он бывает различен. Хондриом хорошо выражен в малодиференцированных клетках, тогда как в высокоразвитых клетках он выступает слабо. В тканях пограничной функции он выражен лучше, чем в тканях внутренней среды.
Обычно хондриом расположен в цитоплазме клеток без какого-либо порядка. Реже (в некоторых разновидностях пограничных тканей) отмечается закономерное расположений хондриома. В них, соответственно гетерополярной диференцировке клеток, хондриозомы располагаются вдоль физиологической оси. В почечном эпителии, в эпителии некоторых выводных протоков слюнных желез (слюнных трубках) базальные части клеток вследствие определенного расположения хондриозом радиально исчерчены. В тканях, клеточные элементы которых аполярны, хондриом, естественно, не имеет такого расположения и распределен в цитоплазме более или менее равномерно.
В генетическом и функциональном отношении хондриом остается еще неизученным. Одни исследователи предполагают развитие хондриозом путем деления, другие — допускают одновременное этим также и новообразование их в цитоплазме, что более вероятно. При размножении клетки хондриом вследствие деления цитоплазмы разделяется между дочерними клетками.
О роли хондриома в жизнедеятельности клетки высказано несколько мнений. Одни исследователи полагают, что с хондриомом связана пластическая функция клетки, проявляющаяся в ходе ее диференцировки.
Примером этого может служить, с одной стороны, образование в цитоплазме тканевых элементов мышечной и нервной систем специализированных структур (миофибрилл и нейрофибрилл), с другой стороны, восприятие клеткой из внешней среды веществ, участвующих во внутриклеточном обмене. В секреторных клетках этот процесс связан с образованием секрета.
Другие исследователи отрицают участие хондриома в пластических реакциях и рассматривают его только как потерявший значение продукт жизнедеятельности клетки. Более правильным будет рассматривать хондриом как весьма изменчивую структуру цитоплазмы, которая при определенных условиях может исчезать или вновь возникать в связи с осуществлением клеткой пластической функции.
Внутренний сетчатый аппарат
Внутренний сетчатый аппарат является постоянным образованием цитоплазмы животных клеток. В растительных клетках он не обнаруживается. Впервые внутренний сетчатый аппарат был обнаружен Гольджи в нервных клетках спинального ганглия (1898). Строение этого органоида не всегда отвечает его названию. Чаще он принимает вид неправильной сети, различно расположенной в цитоплазме. В других случаях его строение имеет характер своеобразного переплета различной толщины балок. Иногда он представлен неправильной формы отдельностями, не связанными между собой и разбросанными в цитоплазме без определенного порядка (рис. 16).
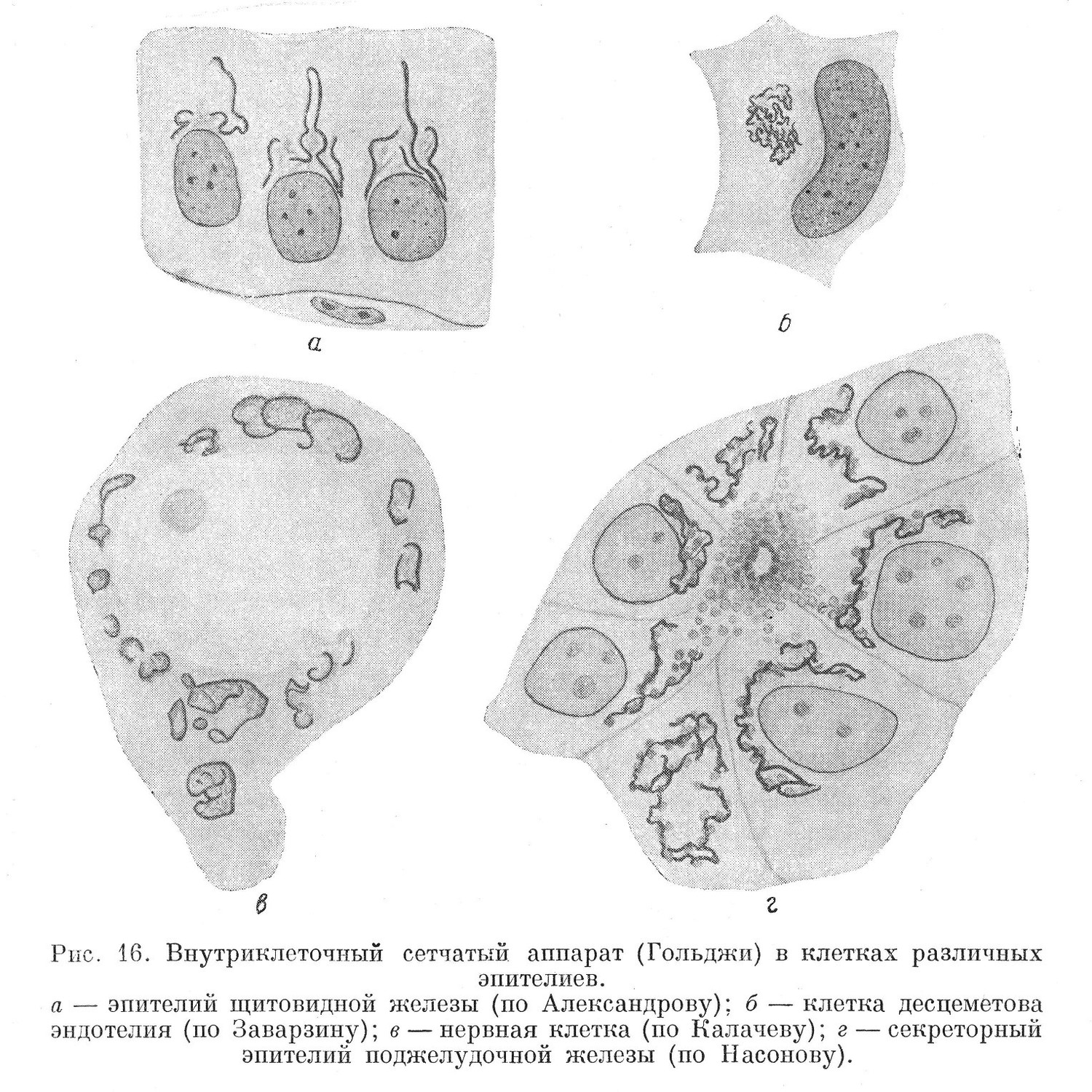
Внутренний сетчатый аппарат, подобно хондриому, весьма чувствителен к изменению внешних условий. При обработке фиксаторами, содержащими кислоту, он разрушается. При выявлении его некоторыми специальными методами исследования одновременно обнаруживается и хондриом. Сетчатый аппарат выявляется при определенном способе обработки ткани азотнокислым серебром, в результате чего на нем выпадает осадок металлического серебра. Такой способ обработки ткани называется импрегнацией. Особенно хорошо удается выявить внутренний сетчатый аппарат клетки, в некоторых случаях одновременно с хондриомом, при осмировании ткани, т. е. при обработке ее четырехокисыо осмия. Здесь также происходит импрегнация, связанная с выпадением металлического осмия.
С помощью осмирования в составе внутреннего сетчатого аппарата удается выявить две составные его части. Одна из них резко окрашивается осмием — осмиоф ильное вещество, имеющее липоидный характер. Другой частью является слабо окрашивающееся осмиофобное вещество белковой природы. Соотношение между липоидным (осмиофиль- ным) и белковым (осмиофобным) веществами в разные периоды жизни клетки изменяется. Если в составе внутреннего сетчатого аппарата белковый компонент доминирует над липоидным, то органоид наиболее устойчив и не разрушается при обработке содержащими кислоту фиксаторами.
В живой клетке внутренний сетчатый аппарат наблюдать не удается. В отличие от хондриома он в живой клетке янусгрюном не окрашивается.
При делении клетки внутренний сетчатый аппарат, подобно хондриому, делится между дочерними клетками.
Расположение внутреннего сетчатого аппарата в клетках разных тканей неоднотипно. Чаще он располагается без заметного порядка в окружности ядра. В пограничных тканях, где клетки полярно диференцированы, он располагается в наружной половине тела клетки, по соседству с ядром. В секреторном эпителии он располагается там, где происходит возникновение секрета. В связи с этим высказано предположение, что внутренний сетчатый аппарат участвует в образовании секрета в клетках. Однако подобным образом он располагается и в других пограничных тканях, которым секреторная деятельность не свойственна.
Представляет интерес изменение положения внутреннего сетчатого аппарата в клетках, образующих эмаль зуба (адамантобласты). Поскольку эти клетки имеют общий источник происхождения с поверхностным эпителием, а следовательно, имеют эпителиальную природу, они рано выявляют гетерополярность. Последнее проявляется также и в том, что внутренний сетчатый аппарат располагается первоначально в наружной половине клетки. Однако непосредственно перед образованием эмали в клетках, образующих эмаль, происходит переориентировка полюсов. То, что было наружным полюсом, становится внутренним, и наоборот, внутренняя (базальная) половина клетки становится наружной. Одновременно происходит инверсия и внутреннего сетчатого аппарата. Последний из наружной половины клетки постепенно перемещается во внутреннюю (рис. 17).
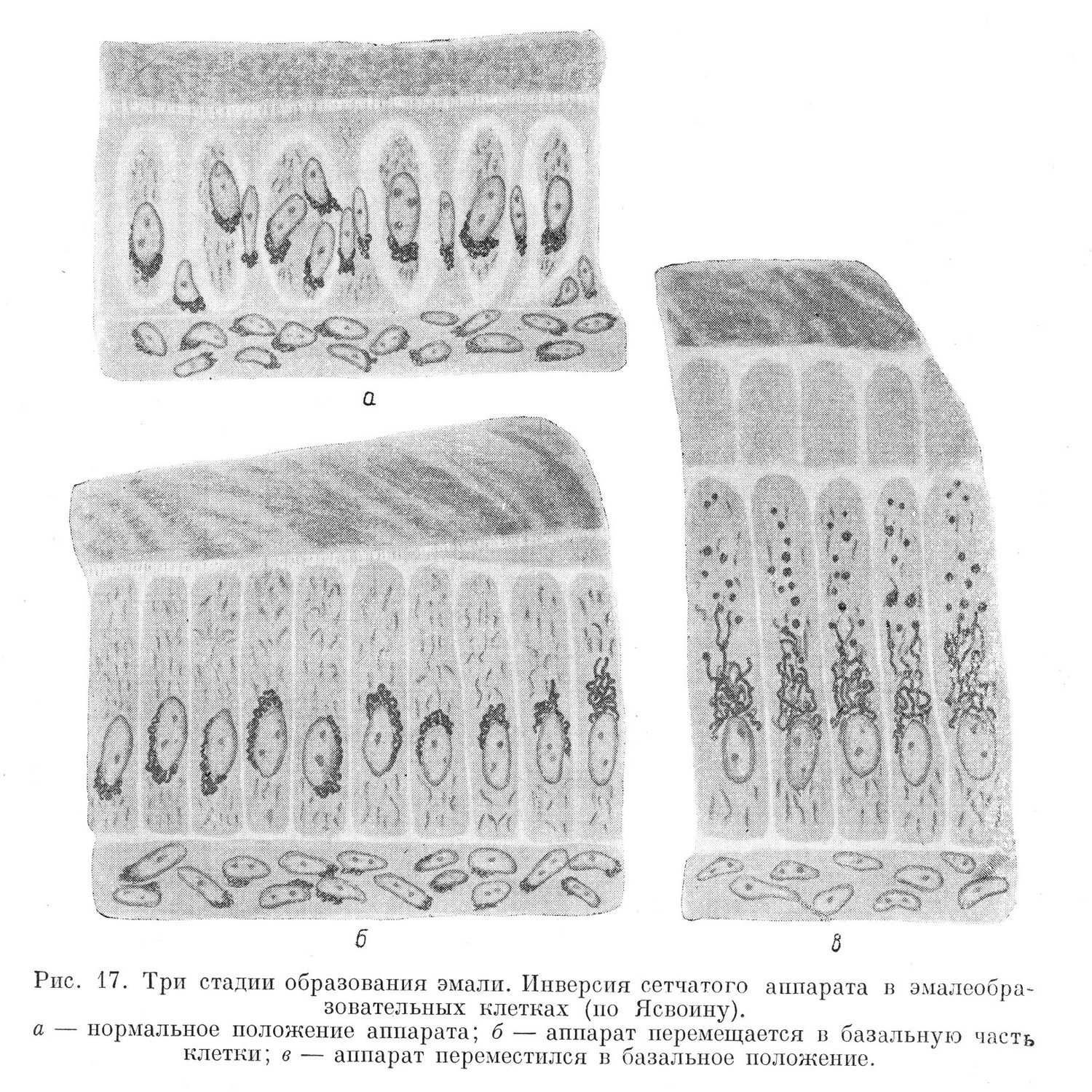
Расположение внутреннего сетчатого аппарата в пограничных тканях в эффектирующей половине клеток, обращенной в сторону внешней среды, является закономерным. Вместе с тем в тканях, клеточные элементы которых аполярны, такая закономерность отсутствует. Здесь чаще всего внутренний сетчатый аппарат располагается в цитоплазме по всей окружности ядра.
Развитие и функция внутреннего сетчатого аппарата клетки не выяснены. Наличие сетчатого аппарата в цитоплазме животных клеток всех 4 типов тканей дает основание предполагать тесную связь его с обменными реакциями клетки. С другой стороны, определенное расположение в пограничных тканях и тканях, клетки которых аполярны, указывает на его отношение, главным образом, к тем обменным реакциям, которые происходят между клеткой и окружающей средой. Этими общими положениями и ограничиваются в настоящее время наши представления о функции внутреннего сетчатого аппарата клеток.
По вопросу о развитии внутреннего сетчатого аппарата можно сделать лишь предположения общего характера, основанные на данных изучения цитоплазмы. Внутренний сетчатый аппарат является, несомненно, диференцированной составной частью цитоплазмы, структура которой обладает большой пластичностью. Это дает основания предполагать, что и строение самого аппарата должно проявлять известную динамичность, обусловленную постоянным обновлением его состава, его новообразованием и распадом. О динамичности его химической структуры свидетельствуют также происходящие в нем изменения соотношения липоидов и белков, обусловленные различным функциональным состоянием клетки и изменениями внешней среды.
Клеточный центр
Клеточный центр был впервые обнаружен в 80-х гг. прошлого столетия. Он был найден в дробящемся яйце морского ежа. Это открытие стоит в прямой связи с описанием митоза в животной клетке, которое было сделано Перемежко в 1879 г. Позднее клеточный центр был описан во многих животных клетках. Лишены его лишь немногие клетки, как, например, зрелая не оплодотворенная яйцевая клетка, абортивные, потерявшие жизнеспособность клетки и др. Клетки растительных организмов, за исключением низших растений, также лишены клеточного центра.
В живой клетке клеточный центр не обнаружен. В клетках различных тканей он располагается неоднотипно. В тканях пограничной функции клеточный центр вместе с внутренним сетчатым аппаратом находится в наружной, эффектирующей половине клетки. Здесь он не всегда занимает центральное положение в соседстве с ядром, а нередко лежит у наружной поверхности клетки. В аполярных клетках клеточный центр занимает центральное положение и оказывает влияние на форму ядра, в той или иной степени оттесняя его к периферии клетки.
Клеточный центр состоит из очень мелких зернышек — центриолей имеющих диаметр от 0,2 до 1 µ (рис. 18). В клетке их бывает одна, чаще две. Значительно реже встречаются клетки с 3 и более центриолями. Гигантские одноядерные и многоядерные клетки содержат до 10 и более центриолей. Центриоли связываются между собой перемычкой — центродесмозой. Центриоли и центродесмоза располагаются в просветленном участке цитоплазмы, который называется сферой или центросферой.
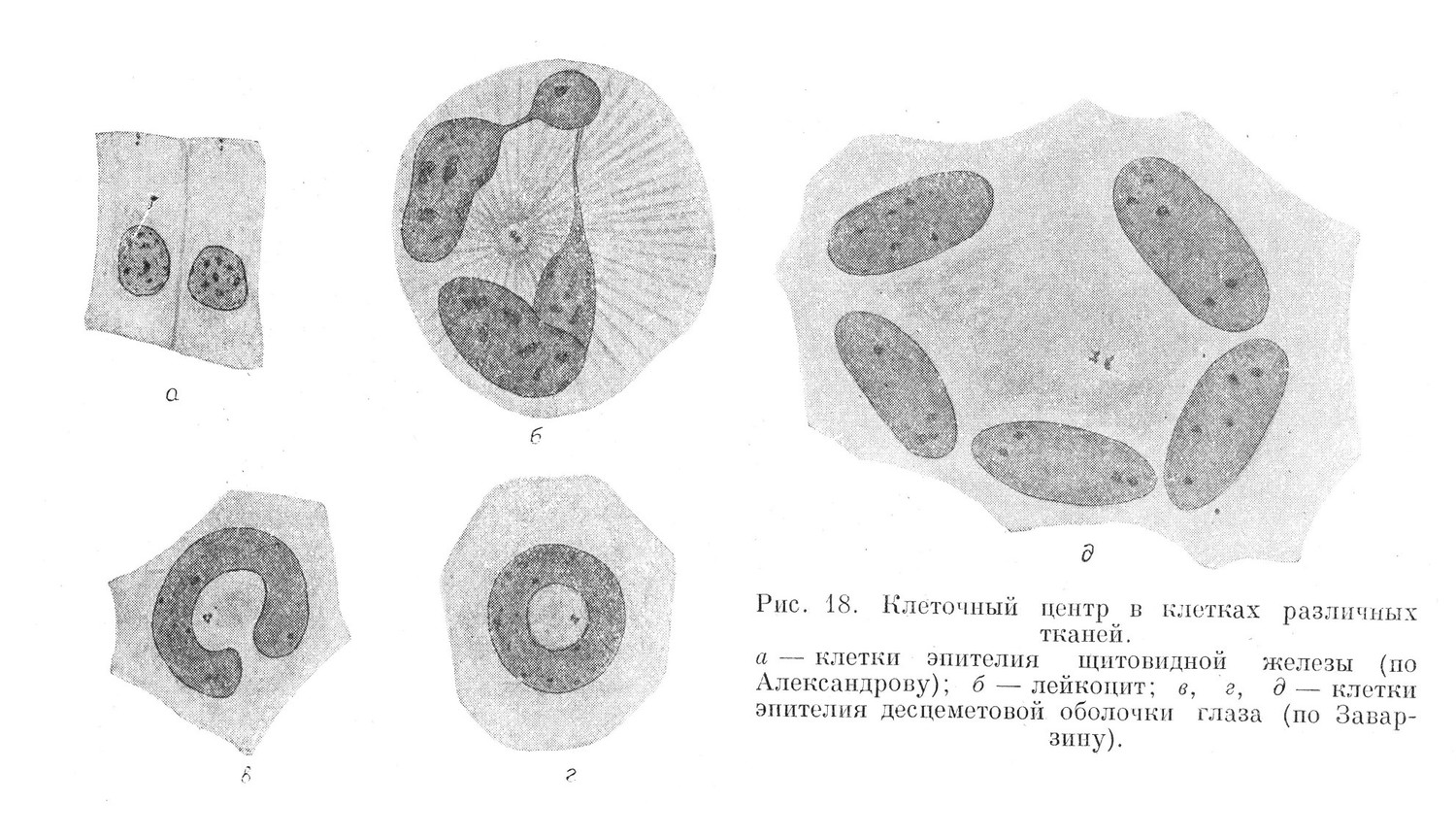
Центриоли, соединенные центродесмозой и расположенные в просветленном участке цитоплазмы, и составляют вместе клеточный центр.
Клеточный центр прижизненно не окрашивается. Это возможно сделать лишь после того, как ткань фиксирована. Он выявляется при окраске железным гематоксилином по методу Гейденгайна. Значительно труднее удается окрасить его сафронином.
В отдельных клетках, преимущественно свободно подвижных (например в лейкоцитах крови), от клеточного центра в цитоплазме проходят радиально расходящиеся тончайшие нити, образующие в целом так называемую лучистую сферу, или астросферу.
Если в цитоплазме имеется зернистость, то она распределяется вдоль этих нитей, и в таких случаях сфера приобретает сложное строение.
Вследствие того, что клеточный центр часто обнаруживается с трудом, высказано предположение о его непостоянстве в цитоплазме клетки. В одни периоды жизни клетки он появляется, в другие — исчезает. Во время митотического деления клетки клеточный центр обнаруживается постоянно, при этом он всегда претерпевает значительные изменения (более подробно об этом будет сказано при рассмотрении вопроса о размножении клеток).
Центриоли способны разделяться. Однако некоторые исследователи допускают возможность образования центриолей не только из предсуществующих — путем их деления, но и в результате новообразования непосредственно из цитоплазмы.
В отношении функции клеточного центра известно лишь то, что он активно участвует в митотическом делении клеток. Однако этим функция клеточного центра, по-видимому, не исчерпывается. Высказано предположение об участии его в развитии локомоторных образований клетки (ресничек, хвостовых нитей, жгутиков). Так, в эпителии, клетки которого имеют на поверхности реснички, в основании каждой реснички находятся мелкие зернышки, называемые базальными тельцами. Последние являются производными клеточного центра, так как произошли вследствие разделения центриолей. В основе хвостовой нити мужской половой клетки (сперматозоида) лежит также клеточный центр. Аналогичные центриолям базальные тельца обнаруживаются в основании жгутиков простейших одноклеточных животных.
В состав клеточного центра, подобно ранее рассмотренным органоидам клетки (хондриому и внутреннему сетчатому аппарату), входят липоиды и белки. Основная часть клеточного центра — центриоли являются наиболее плотными частями цитоплазмы. В пользу этого говорят некоторые косвенные данные, а именно, более медленная потеря краски при вытягивании ее из препарата по сравнению с остальной цитоплазмой.
Все изложенное дает основания рассматривать клеточный центр как высокодиференцированную изменчивую часть цитоплазмы, связанную с функцией митотического деления клетки и развитием в ней локомоторных образований.
Включения
Жизнедеятельность клетки характеризуется непрерывно совершающимся обменом веществ. В связи с обменом в клетке периодически накапливаются и исчезают продукты различного состава. Одни из них находятся в диффузном состоянии и при малом их количестве микроскопически не обнаруживаются. Другие образуют в клетке микроскопически хорошо видимые скопления, которые имеют вид вакуолей, если вещество находится в жидком состоянии, или гранул, если оно плотной консистенции, или, наконец, кристаллов, в случае его способности в определенных условиях к кристаллизации. Все эти продукты носят название включений.
Включения являются временными образованиями в клетке и в ее жизнедеятельности имеют различное значение. Одни из них в процессе внутриклеточного обмена в том или ином виде входят в состав живого вещества, т. е. ассимилируются данной или другой клеткой. Они составляют группу трофических включений. Другие являются продуктами диссимиляции. Они временно пребывают в клетке и выделяются ею в окружающую среду. Их называют экскреторными включениями. Имеются включения, постоянное образование которых и выделение во внешнюю среду является главной функцией клетки. К ним относятся секреторные включения. Наконец, в Цитоплазме клеток встречаются продукты, связанные с другими различными сторонами жизнедеятельности клеток и обычно представленные в виде мелких гранул или вакуолей. Их объединяют в группу неспецифических включений.
Итак, включения не представляют диференцированной части живого вещества — цитоплазмы и кариоплазмы, а являются лишь продуктами, связанными с жизнедеятельностью клеток. В большинстве случаев они содержатся в клетке в течение короткого промежутка времени.
Трофические включения. Сюда относятся, главным образом, жир, гликоген и липоиды. Накопление их в цитоплазме происходит при активном участии самих клеток из продуктов, которые приносятся к тканям кровью и лимфой или непосредственно воспринимаются из внешней среды, или, наконец, являются продуктами внутреннего обмена самой клетки.
Жировые включения обнаруживаются в клетках почти всех тканей организма. Чаще они встречаются в небольшом количестве в виде мелких капелек. Подобные жировые включения следует рассматривать как проявление внутриклеточного обмена веществ. Их можно встретить в цитоплазме клеток различных разновидностей соединительной и эпителиальной тканей, а также в тканевых элементах мышечной и нервной систем. При голодании организма эти жировые включения не исчезают, поэтому их называют постоянным жиром. В клетках отдельных разновидностей тканей внутренней среды и пограничных тканей в качестве энергетического запаса накапливается большое количество жира, но при голодании организма ткани теряют его;, такой жир называется расходным. Подобные жировые включения обнаруживаются в клетках костного мозга, в печени.
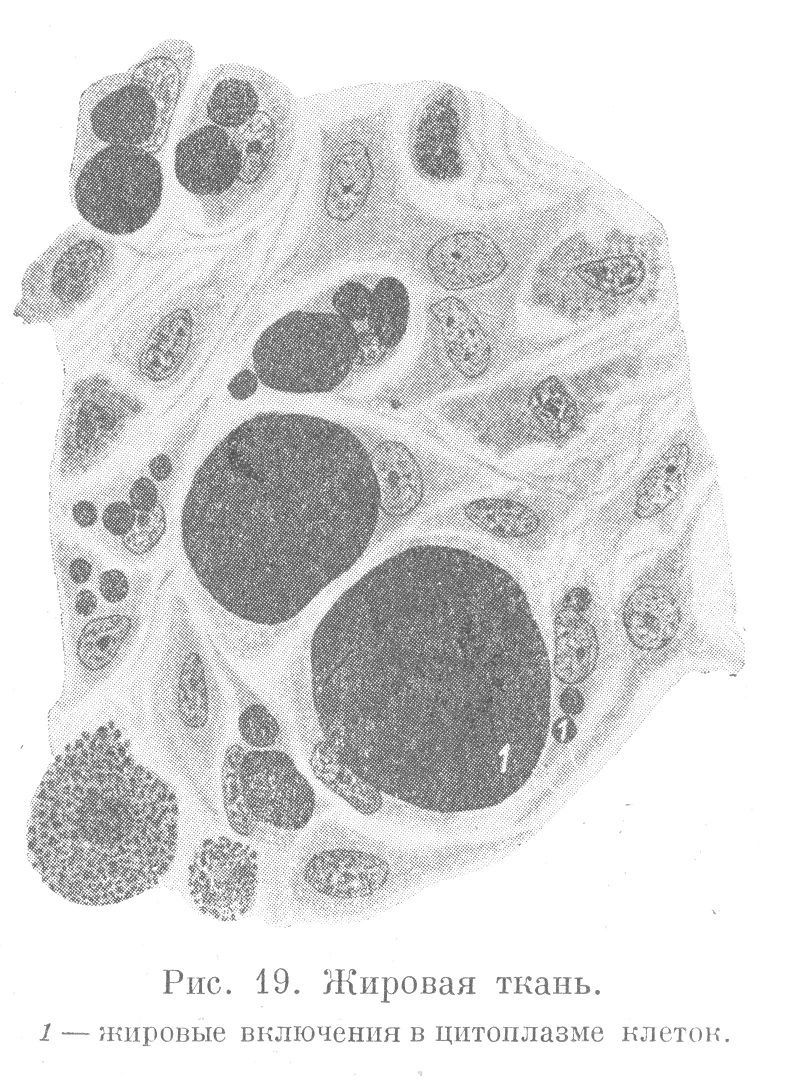
Особенно много жировых включений в некоторых отделах рыхлой неоформленной соединительной ткани, которая в связи с этим называется жировой тканью. Клетки соединительнотканной природы, содержащие жир, называются жировыми клетками. Каждая из них содержит в цитоплазме относительно крупную каплю жира, вследствие чего принимает шарообразную форму. В таких клетках цитоплазма, содержащая ядро, окружает жировую каплю в виде узкого ободка (рис. 19). Жировые включения хорошо выявляются при обработке ткани четырехокисыо осмия, при окраске Суданом III и шарлахротом.
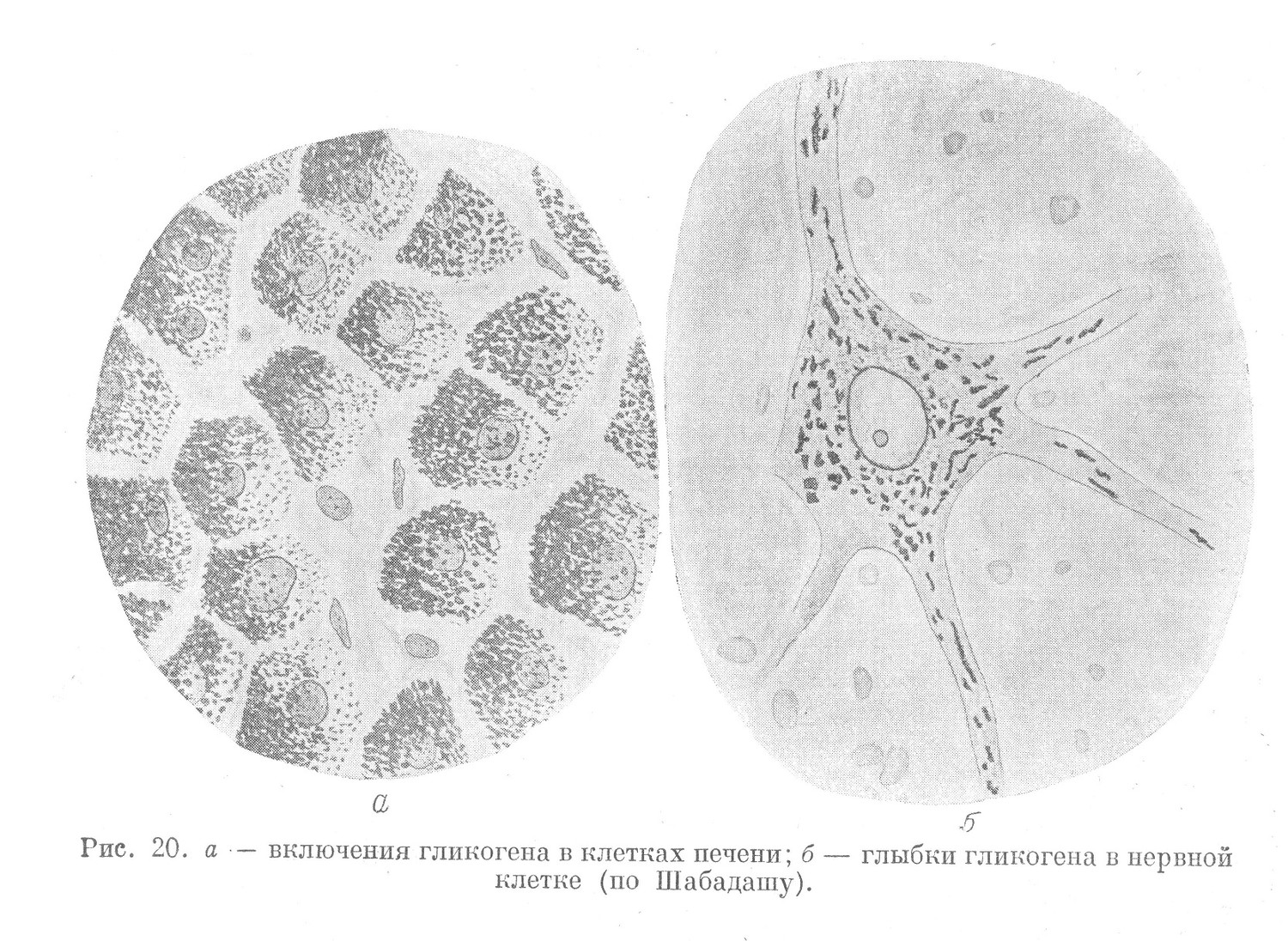
Включения гликогена, также как и жир, являются энергетическим материалом. В небольшом количестве гликоген встречается во всех клетках организма и связан с их обменом. Более значительные скопления в качестве запасного питательного материала встречаются в сравнительно немногих тканях, например в печени, поперечно-полосатой мускулатуре и др. Гликоген находится в клетках в виде различного размера глыбок неопределенного очертания (рис. 20). Гликоген растворим в воде и нерастворим в крепком спирте, который и применяют в качестве фиксатора. Выявляется гликоген при окрашивании кармином Беста и другими методами.
Липоидные, или жироподобные, вещества обнаруживаются во всех клетках. Они образуют преимущественно рыхлые соединения с белками и входят в состав цитоплазмы живого вещества. Однако в клетках отдельных тканей они представлены в виде обособленных от цитоплазмы скоплений и могут быть названы включениями. Такого рода липоидные включения находятся в некоторых вспомогательных структурах нервной системы (например миэлин в шваннодских клетках) или в виде гранул в клетках коры надпочечника, в клетках желтого тела и других элементах. Липоидные включения возникают в клетке, невидимому, за счет доставляемых веществ.
Экскреторные включения представляют продукты распада (диссимиляции) части живого вещества клетки. Они обособляются в клетке, после чего выводятся в окружающую среду. К ним относятся мочекислые соли, мочевина, желчные пигменты и др.
Пигментные включения имеют различный состав и разное значение в жизнедеятельности клетки. Часть из них участвует в газовом обмене. Сюда относится гемоглобин, сложное соединение белкового характера, содержащее железо. Гемоглобин в большом количестве содержится в составе эритроцитов, диффузно пронизывая их цитоплазму. Другие пигменты не участвуют в дыхательной функции, а придают ткани и клетке ту или иную окраску. К ним относятся меланины, липохромы, липофусцины, гемосидерин и гематоидин.
Последние два пигмента являются продуктами распада дыхательного пигмента.
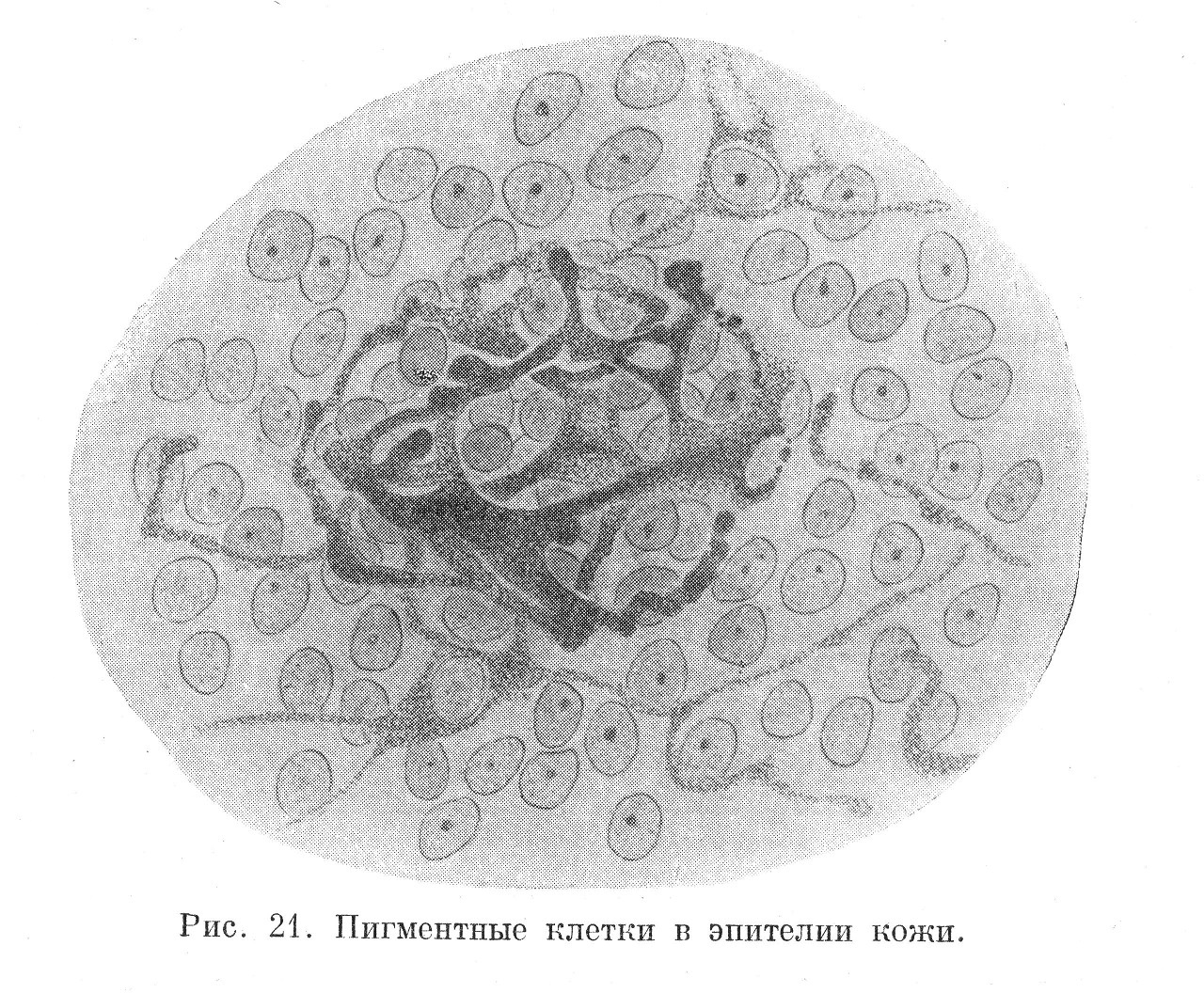
Меланины — это черные и бурые пигменты. Находятся они в хроматофорах — клетках рыхлой соединительной ткани (рис. 21), в пигментной соединительной ткани, в сосудистой оболочке глаза и в клетках пигментного эпителия сетчатки. Меланины обнаруживаются также в клетках эпителия и соединительной ткани кожи при ее загаре.
Липохромы — пигменты желтого и красного цвета. В цитоплазме они расположены в виде мелких капелек жироподобного вещества. Липохромы обнаруживаются в лютеиновых клетках (желтого тела), в жировых клетках, в сетчатке и других образованиях.
Липофусцины в цитоплазме располагаются в виде мелких зернышек и весьма близки к липохромам. Встречаются в, клетках печени, почек, надпочечных желез и других органов.
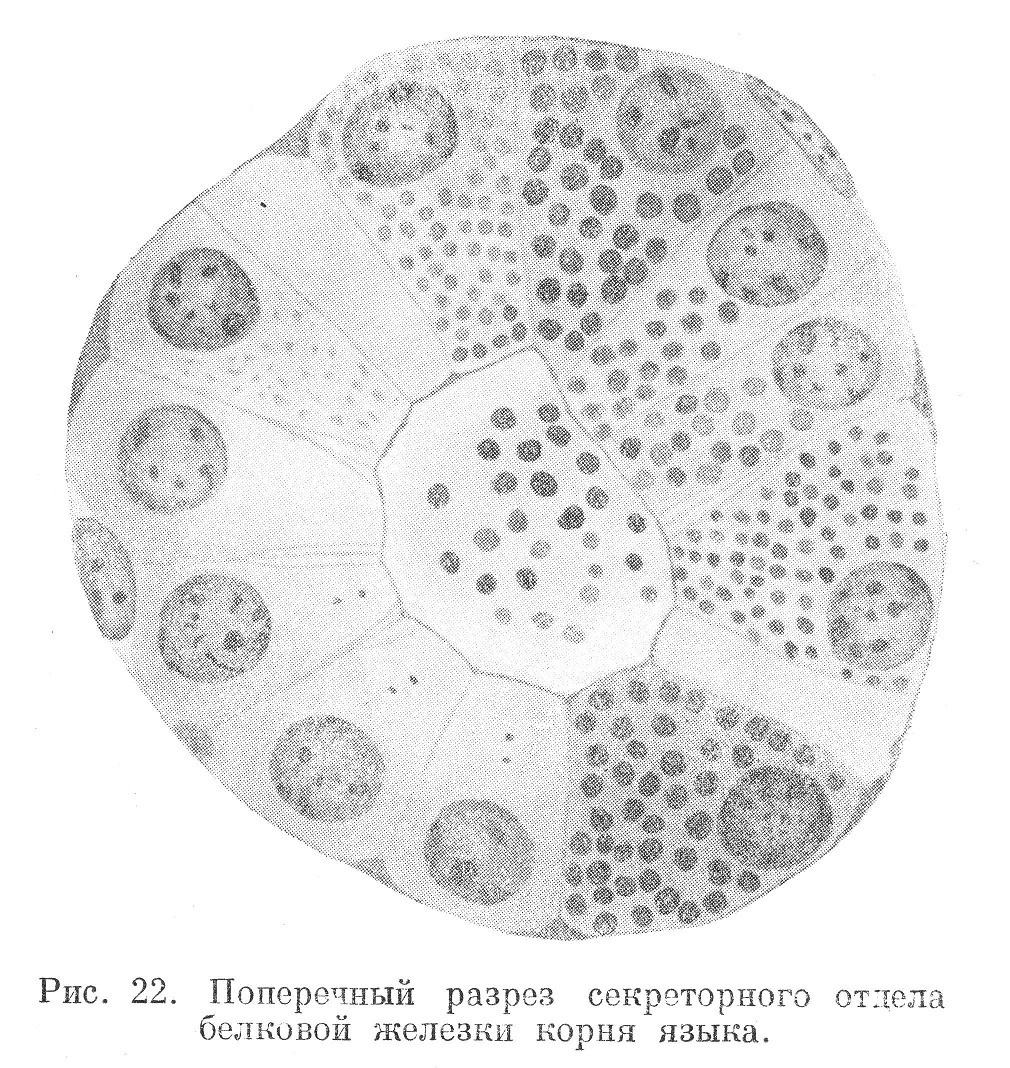
Секреторные включения представляют разнообразную группу образований, вырабатываемых в цитоплазме клеток. Они называются секретом и имеют различное значение в организме. Секрет возникает в клетках экзокриновых желез и выделяется во внешнюю среду (рис. 22). Агрегатное состояние секрета различно. В одних случаях он бывает жидким и представлен различного размера вакуолями. В других — он более плотный, в виде зерен или гранул. Более подробно о развитии и строении секрета будет сказано при рассмотрении эпителиальных тканей и, в частности, желез.
Неспецифические включения имеют различный вид и строение. В большинстве случаев — это мелкие капельки или гранулы, которые то появляются, то исчезают. Иногда в клетках образуются более крупные вакуоли, содержащие растворы солей или жидкость, имеющую белковый, липоидный и углеводный состав. Появление неспецифических включений нередко связано с нарушением обмена веществ в клерке.
Метаплазматические образования и межклеточные (промежуточные) вещества
Под именем метаплазматических образований и промежуточных веществ тканей понимают структуры, возникшие в процессе развития и диференцировки клеточных элементов. Они тесно связаны с жизнедеятельностью клеток и появляются в их онтогенезе, протекающем в составе ткани. Метаплазматические образования находятся в цитоплазме клеток. К ним относятся тонофибриллы эпителиальных тканей, миофибриллы мышечных клеток и мышечных волокон и нейрофибриллы нервных клеток. Промежуточные вещества лежат между клетками и находятся в тесной связи с определенными клеточными формами. Они образуют не имеющее клеточного строения живое вещество. Последнее представлено аморфной бесструктурной массой, преколлагеновыми и коллагеновыми фибриллами, эластическими волокнами и мембранами. Метаплазматические образования представляют высокодиференнированные составные части клеток. Промежуточные вещества родственны клеткам, но морфологически связаны не со всеми клеточными элементами ткани.
В функциональном, морфологическом и генетическом отношении метаплазматические образования следует представлять как специализированные составные части цитоплазмы. Они возникают в клетках в процессе развития тканей. В клетках эмбриональных зачатков они отсутствуют.
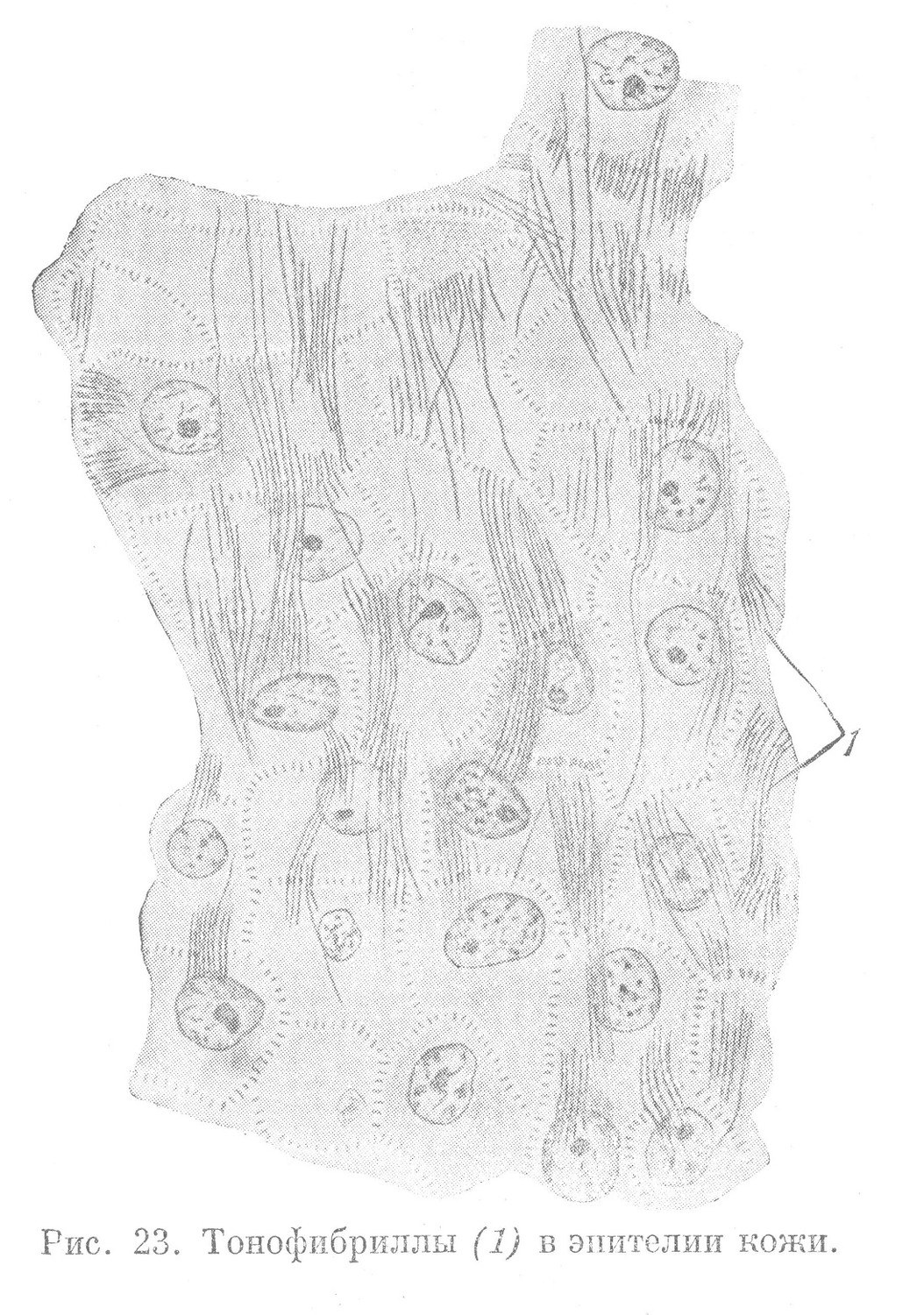
В эпителиальных тканях метаплазматические образования представлены тонкими, длинными фибриллами, образующими пучки и выполняющими опорную функцию. Проходя в определенных направлениях через многие клетки эпителиального пласта, они пересекают межклеточные щели, располагаясь в межклеточных мостиках (рис. 23). В некоторых разновидностях эпителия тонофибриллы обнаруживаются лишь в цитоплазме отдельных клеток и за их пределы не выходят. Такие клетки в составе эпителиального пласта выполняют опорную функцию.
В тканевых элементах мышечной системы метаплазматические структуры представлены миофибриллами. Они имеют вид тонких волоконец, обусловливающих функцию сокращения (рис. 24). В клетках гладкой мышечной ткани миофибриллы на всем протяжении однородны. Они свободно лежат в цитоплазме и идут параллельно друг другу вдоль тела клетки.
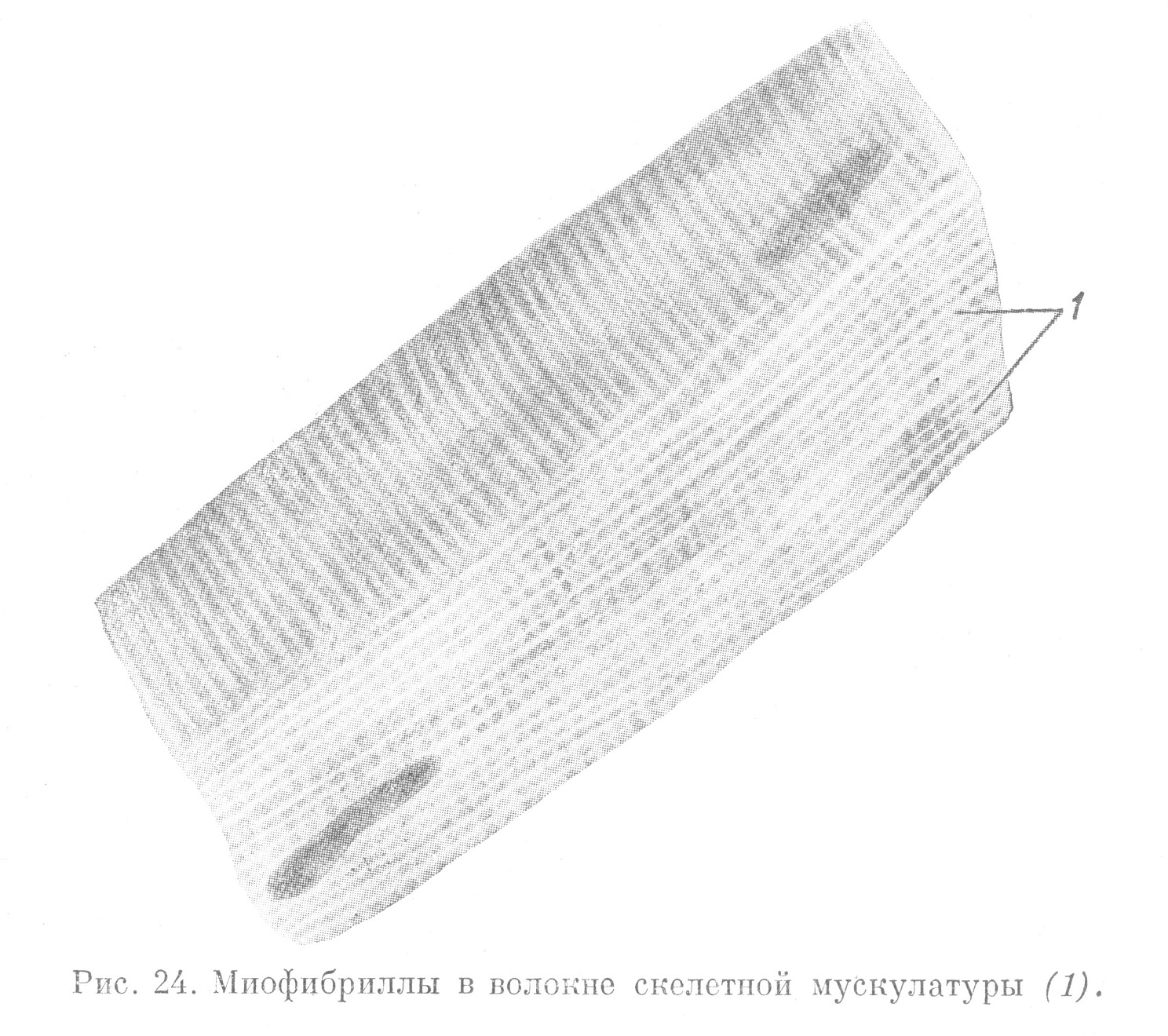
В мышечных волокнах скелетной мускулатуры все миофибриллы между собой связаны и образуют в каждом мышечном волокне единый сократимый аппарат. Строение миофибрилл неоднородно. В составе их находится вещество, образующее диски, которые, чередуется по ходу миофибрилл. Ввиду того, что различные диски обладают неодинаковым коэфициентом преломления, одни из них кажутся светлыми, другие — темными. Это придает миофибриллам, а вместе с тем и всему мышечному волокну поперечную исчерченность. Отсюда и происходит название мускулатуры — поперечно-полосатая (рис. 24).
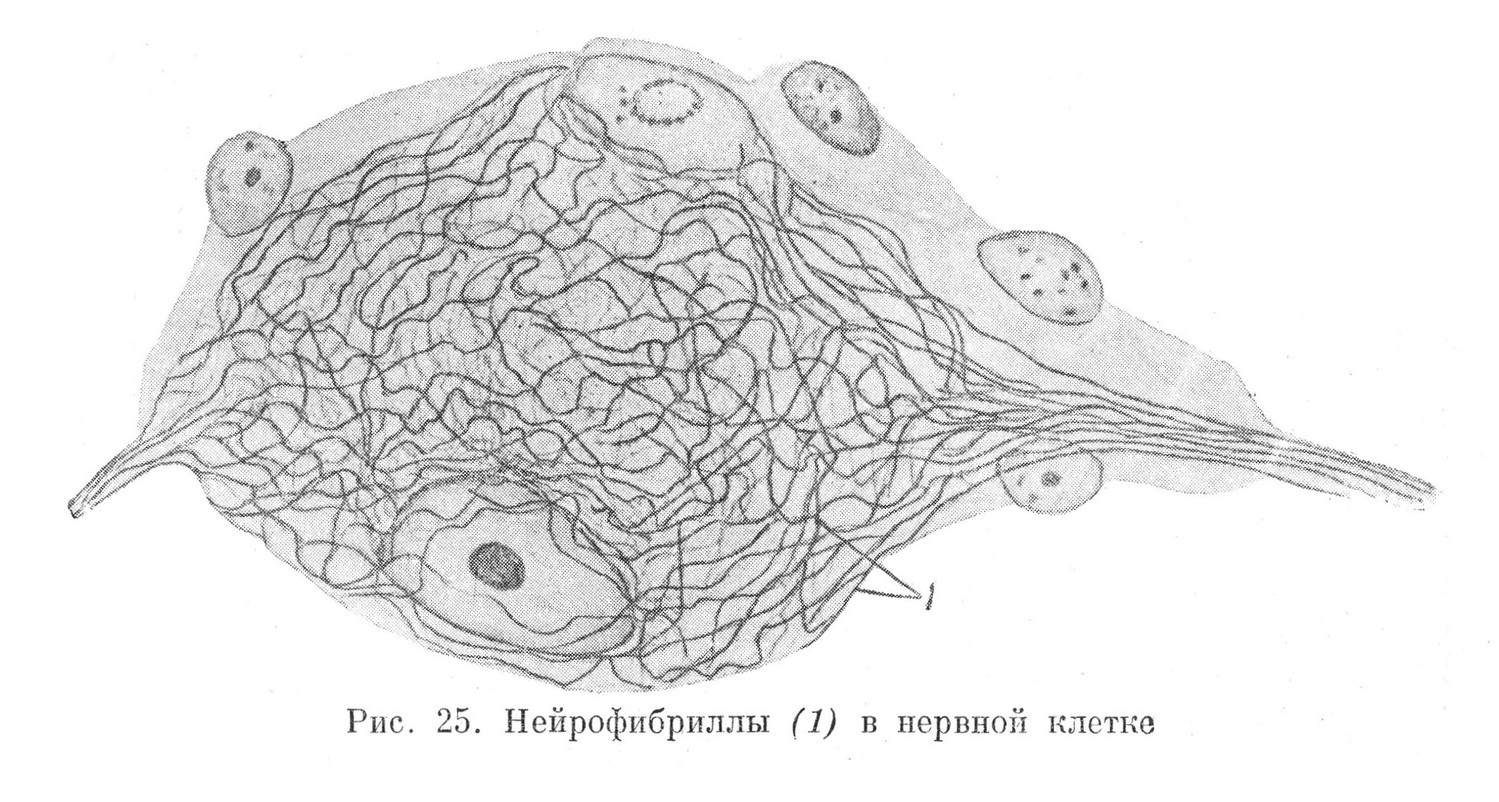
Метаплазматические образования нервных клеток,— нейнофибриллы однородны и проходят как в теле, так и в отростках нервных клеток, на всем их протяжении. За пределы невронов они не выходят (рис. 25). Вместе с цитоплазмой (нейроплазмой) нейрофибриллы проводят нервный импульс.
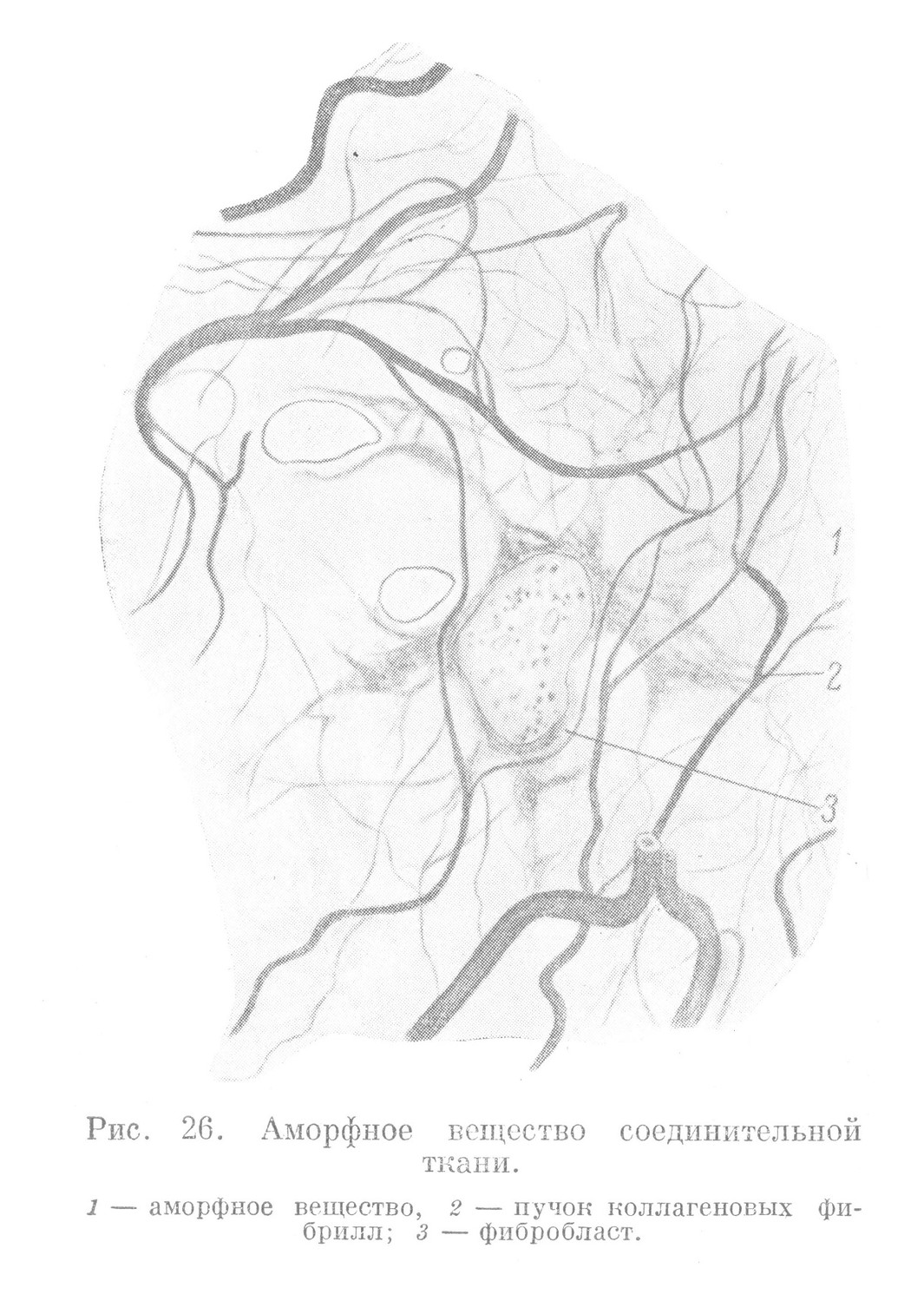
Промежуточные вещества выполняют механическую, опорную функцию тканей. Они хорошо развиты в тканях внутренней среды, особенно в тех разновидностях ее, которые входят в состав скелета. В некоторых опорных тканях (в костной ткани) промежуточное вещество находится в связи с неорганическими соединениями (различными солями), вследствие чего оно приобретает значительную твердость и прочность. Наиболее распространенной частью промежуточного вещества являются находящиеся на разных стадиях развития коллагеновые фибриллы. Наиболее молодые, менее развитые из них, называются преколлагеновыми. Они возникают в тканях постоянно на протяжении всей их жизни. Их можно найти в большом количестве во многих тканях внутренней среды. Их называют аргирофильными фибриллами, так как они хорошо выявляются при обработке ткани солями серебра (рис. 26). Аналогичные фибриллы ретикулярной ткани называют ретикулиновыми волокнами. Расположение указанных фибрилл часто представляется беспорядочным, и нередко они имеют вид неправильной сети.
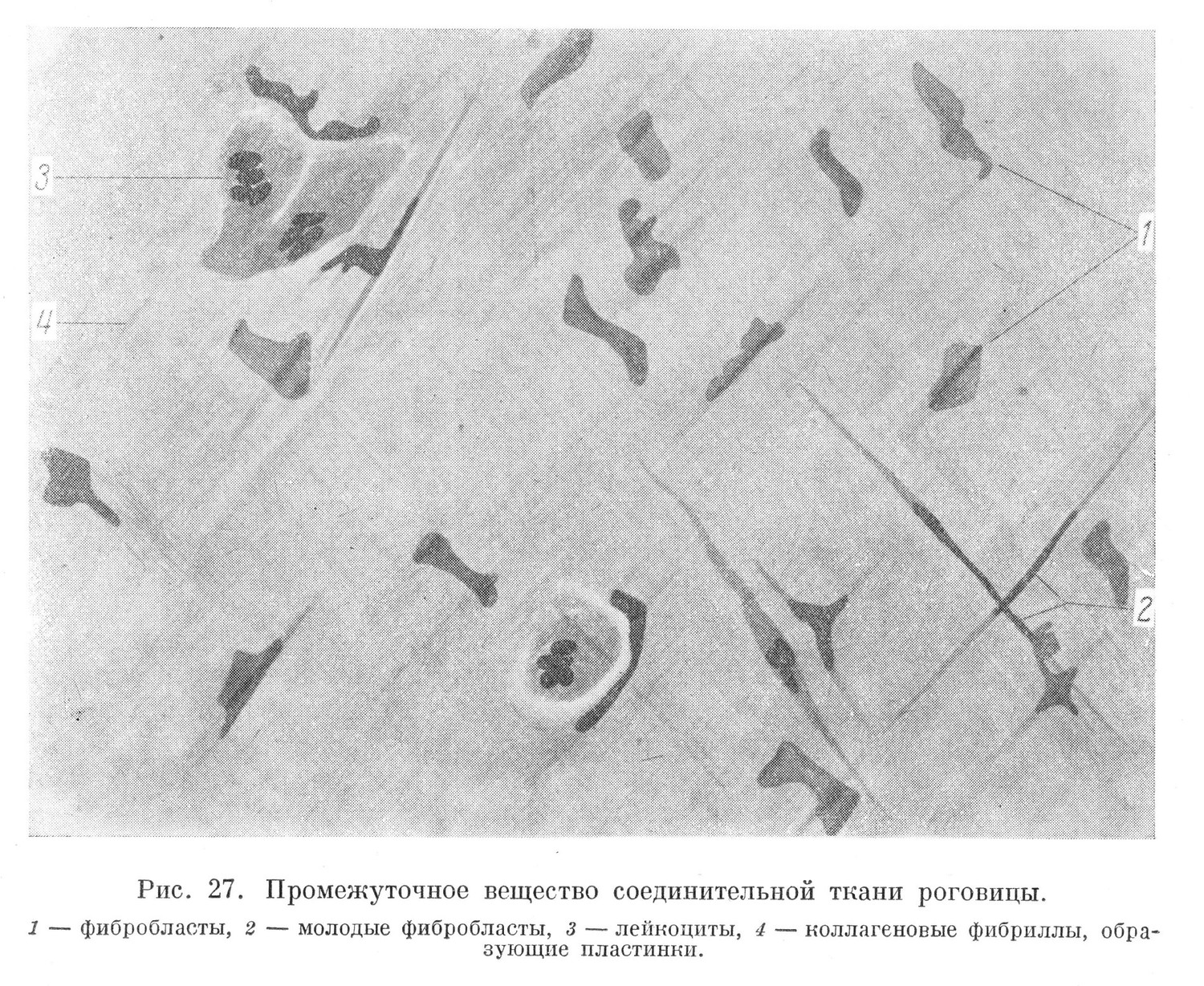
Коллагеновые фибриллы в различных тканях располагаются неоднотипно. В одних случаях они образуют различной толщины пучки, в которых фибриллы спаяны между собой аморфным веществом. Пучки проходят или в беспорядке (рыхлая и плотная неоформленная соединительная ткань), или строго закономерно (плотная оформленная соединительная ткань) (рис. 27). В других тканях из параллельно идущих коллагеновых фибрилл, спаянных аморфным-веществом, образуются пластинки, наложенные одна на другую (собственное вещество роговицы, пластинчатая костная ткань) (рис. 27). Фибриллы хрящевой ткани, аналогичные коллагеновым, называют хондриновыми, а костной ткани — оссеиновыми.
Аморфное вещество образует либо пластинки, соединенные между собой под острым углом (например рыхлая неоформленная соединительная ткань), либо отложения между клетками, заключающие в себе коллагеновые фибриллы, как это имеет место, например, в хрящевой или костной тканях. Это бесструктурное вещество во многих случаях переходит непосредственно в наружную часть цитоплазмы оседлых клеток (фибробластов или других аналогичных им клеток).
Эластические волокна имеют различную толщину и по своему ходу могут разветвляться. Часто они образуют различной плотности сети. В некоторых органах (гладкая мышечная ткань кровеносных сосудов) образуются эластические мембраны. Эластические волокна и мембраны обладают свойством растягиваться и снова возвращаться в прежнее состояние. Они встречаются в тех местах, где степень растяжения тканей постоянно изменяется, например в стенке многих кровеносных сосудов, в стенке дыхательных путей и легких, на некоторых участках кожи и т. и.
Образование промежуточного вещества находится в прямой связи с развитием ткани, с диференцировкой ее клеточных элементов. Высказаны две теории, объясняющие этот процесс. Одна из них является наиболее распространенной и в отношении многих тканей вполне доказана. Ее можно назвать теорией эктоплазматического образования промежуточного вещества. Суть этой теории заключается в том, что цитоплазма оседлых клеток разделяется на две части: наружную — эктоплазму и внутреннюю — эндоплазму, которые различаются между собой. Эктоплазма по сравнению с эндоплазмой более плотна и лишена органоидов и включений. Между ними нет резкой границы, а чаще отмечается постепенный переход.
В ходе развития ткани в эктоплазме указанных клеток появляются тончайшие фибриллы промежуточного вещества, которые представляют преколлагеновые или молодые эластические волоконца. Одновременно с этим возникает аморфное вещество. Таким образом, источником образования промежуточного вещества является эктоплазма клеток, которая в определенных условиях претерпевает метаморфоз. Восстановление эктоплазмы в свою очередь происходит со стороны эндоплазмы.
Другое мнение об образовании промежуточного вещества выражено в так называемой секреционной теории. Сторонники секреционной теории исходят из того, что клеточным элементам тканей, где образуется промежуточное вещество, независимо от их природы свойственна секреционная деятельность. Выделившийся в межклеточные пространства секрет превращается в различные формы промежуточного вещества. Слабым местом этой теории является прежде всего то, что если секреторная деятельность ясно проявляется пограничными тканями, то способность к ней клеток тканей внутренней среды совершенно не доказана. Нет достаточно убедительных морфологических данных, показывающих выделение какого-либо секрета указанными тканями в межклеточные щели, и не доказано превращение его в промежуточное вещество.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 