Существует мнение, что морфологический метод уже полностью использован в гематологии, что этот метод вряд ли может внести что-либо новое при изучении болезненного процесса. Однако с этим мнением нельзя согласиться хотя бы уже потому, что наши сведения о клеточных структурах обычно ограничиваются изучением процессов по фиксированным отдельным моментам процесса; наши же стремления направлены на изучение живой клеточной структуры, рождающейся в цикле разнообразных жизненных процессов, развивающейся и изменяющей свою форму под влиянием их динамического воздействия.
По воззрениям большинства современных исследователей клетка является простейшей, элементарной биологической системой. Отдельные авторы (Альтман, Гайденгайн (Altmann, Heidenhain) и др.) считают, что способностью самостоятельно жить обладают также некоторые структуры клетки, митохондрии; однако это еще до сих пор не доказано, точно так же как не доказана теория Гука (Hueck), что и межклеточное вещество является живой системой.
Форменные элементы крови — эритроциты и лейкоциты — представляют собой наиболее благодарный объект для изучения основ структуры и функции живого многоклеточного организма, так как они являются частью живыми, частью отмирающими структурами, эксперимент над которыми приближается к эксперименту над одноклеточными организмами — протозоа. Поэтому красные кровяные тельца служат у биохимиков излюбленным материалом для изучения главным образом проницаемости мембран.
Морфологическое изучение кровяных телец состоит: 1) из наблюдения и эксперимента в живом неокрашенном виде и 2) из изучения окрашенных препаратов. Последний метод является микрохимическим методом для выявления клеточных структур, химизма этих структур и, наконец, для выяснения внутриклеточных процессов. Как осторожны мы должны быть с оценкой внутриклеточных реакций, прекрасно иллюстрирует в своей работе Дейтш (Deutsch). Он показал, что с помощью краски-индикатора не всегда удается установить изменение концентрации водородных ионов внутри клетки, так как благодаря множеству поверхностей, входящих в состав клеточного вещества, адсорбированная на них краска имеет меньшую электрическую диссоциацию, чем краска примененного раствора; поэтому, несмотря на то, что реакция среды не изменилась, краска-индикатор изменяет свой цвет. Дейтш иллюстрировал это на растворе краски тимолблау и эмульсии из бензола и воды. Согласно теории Оствальда (Ostwald) об индикаторах, ионы тимолблау (диссоциированные молекулы) желтого цвета, а недиссоциированные молекулы — красновато-фиолетовые; в опыте Дейтша пена раствора краски и эмульсии из бензола и воды окрашивалась в красновато-фиолетовый цвет (вследствие наличия многих поверхностей, на которых адсорбировалась краска, и образования недиеооциированных молекул), тогда как раствор этой же краски был желтым.
На этом примере мы видим, что при образовании множества соприкасающихся поверхностей краска-индикатор, адсорбируясь на поверхностях, резко изменяет свою окраску и симулирует таким образом изменение реакции среды. Это, конечно, только одна из многих возможностей неправильного толкования при окраске клеток.
Так же велики трудности, когда приходится решать вопрос о проницаемости. оболочки клетки для тех или иных красочных веществ или реактивов. Полупроницаемость животных мембран (проходимость их для воды и непроходимость для растворенных веществ) при различных физиологических процессах может изменяться; так, оболочка клетки при определенном физиологическом состоянии становится проходимой для сахара, аминокислот и солей. Чтобы объяснить такую изменчивость клеточной проницаемости, Гёбер (Hober) ввел понятие о «физиологической» проницаемости в отличие от физической. Физиологическая проницаемость обусловливается изменчивостью оболочки под влиянием внутриклеточных процессов; это изменчивость оболочки клетки носит по Гёберу регуляторный характер. Изменяется ли при этом только состав отдельных участков оболочки или же варьирует и величина пор между ее составными частями [Руланд (Ruhland)], так что большие молекулы механически получают доступ к клетке, пока еще твердо не установлено. С физиологической проницаемостью клетки мы должны будем считаться при применении целого ряда прижизненных окрасок.
Работы Варбурга последних лет доказывают, что теория наркоза Майера-Овертона построена на ложных началах—наркотическое действие вещества на клетку обусловливается его способностью адсорбироваться на поверхности и связывать дыхательные ферменты. Здесь выявился случайный параллелизм между коэффициентом распределения этих веществ между водой и липоидами (Майер-Овертон) и их адсорбиционными свойствами. Интересные данные по этому поводу приводит Баадер (Baader). Он указывает на значительную разницу между растворимостью чужеродных веществ в воде и в крови; так, например, коэффициент растворимости, т. е. цифровой показатель растворимости, для хлорированных углеводородов (хлороформ, четыреххлорисрый углерод, трихлорэтилен) дает нарастающие цифры растворимости в следующем порядке: вода, физиологический раствор поваренной соли, сыворотка крови, кашица из эритроцитов; при этом различие в растворимости между водой и кровью может достигать 100% [Люче (Luce)]. Не учтя этого различия, в производство был введен как заместитель бензина трихлорэтилен (считавшийся безвредным веществом), давший сотни отравлений и много смертных случаев. При испытании растворимости эфира в крови, воде и мозговой кашице оказалось, что она во всех трех случаях одинакова, т. е. что коэффициент распределения эфира между кровью и мозгом равен 1:1. Это звучит парадоксально, если учесть высокую растворимость жира эфиром и богатство мозга липоидами.
Что касается так называемой осмотической резистентности эритроцитов, то здесь приходится считаться с фактором, который на первый взгляд также кажется парадоксальным: в живой протоплазме сохраняются то большие, то меньшие концентрации электролитов, чем в окружающей среде, и только при отмирании клетки происходит выравнивание концентраций. Ирвин (Irwin) доказал это на водорослях, в которых было установлено 0,128 мг свободных растворенных хлоридов, тогда как в окружающей воде их почти совсем не было. Все исследования последнего времени показывают, что концентрация водородных ионов внутри клеток иная, чем в окружающей их среде. Это относится и к кровяным тельцам. Доказательство этого мы находим у асцидий. Плазма крови этих животных имеет слегка кислую реакцию (pH = 6,69), но в плазме плавают зрелые кровяные тельца с pH от 2 до 3! Кажется совершенно невозможным понять, как маленькие тельца, имеющие огромную поверхность обмена с окружающей жидкой средой, могут поддерживать такую разницу ионной концентрации. Из этого примера ясно видно, что при работе с живой материей мы встречаемся со многими неизвестными, причем их количество значительно увеличивается по сравнению даже с самыми сложными структурами мертвой ткани. Также не известны нам те коррективы, которые должны быть введены при переводе данных, полученных при исследовании отмирающих или умерших систем, на их предшествовавшее живое состояние. Бауер (Bauer) утверждает, что при отмирании устанавливается равновесие между живой системой и окружающей средой и что живая система, пока она жива, никогда не приходит в состояние равновесия с окружающей средой, что является результатом функции ее структур.
По Нернсту (Nernst), внутри живой системы существует состояние так называемого динамического равновесия, при котором абсолютного равновесия никогда не наступает, но «уравновешивается взаимный обмен». Однако, к каким бы выводам ни пришла научная мысль относительно определенных закономерностей в течении жизненных процессов, в них должен найти отражение принцип уравновешенности процессов, происходящих внутри живого работоспособного организма; этот принцип выражается в различных константах — концентрации водородных ионов, координации гормональных взаимоотношений и т. д. Уравновешенность замкнутой работоспособной системы резко выступает при анализе живого организма наряду с также резко выступающим неравновесием его, живого организма, в отношении окружающей среды. Нужно думать, что в единстве противоположностей внутри и вне организма протекающих процессов заключается сущность раздражимости — основного свойства живого организма.
Постоянство внутренней среды у высших животных достигает очень тонких установок. Так, для человека пределы колебаний концентрации водородных ионов в плазме крови, совместимые с жизнью, составляют pH = 7,0-7,8, т. е. по грубому подсчету между 1 и 5 г ионов водорода в 108 литрах. Излишне доказывать, говорит Баркрофт, незначительность абсолютного количества этой величины. Распределение 1—5 г водородных ионов в 108 литрах плазмы соответствует распределению 1—5 г ионов водорода в плазме крови половины населения Соединенных Штатов Америки. Колебания таких минимальных концентраций достаточны, чтобы человек из состояния полного здоровья перешел в состояние предсмертных конвульсий. Важную роль в буферной функции крови играет гемоглобин. Когда кислота прибавляется к плазме крови, она (или ее эквивалент) переходит в эритроциты (проницаемые для кислот) и там нейтрализуется системой бикарбонат-гемоглобин, благодаря чему реакция плазмы остается сравнительно неизменной. Кроме буферного действия жидкости и соприкасающихся с ней тканей, в удержании колебаний концентрации водородных ионов в определенных пределах участвуют почки (выделение кислоты, удержание щелочей и продукция аммиака), а также легкие (выделение кислоты). К этому присоединяется еще наиболее тонкая регуляция посредством нервной системы (дыхательный центр). Функция нервной системы наиболее ранима при нарушении констант также в смысле постоянства температуры, содержания воды, сахара, кислорода, кальция и т. д. «Предполагать высокое интеллектуальное развитие, — говорит Баркрофт, — в среде, свойства которой не стабилизированы, — это значит искать музыку в треске плохой радиопередачи или зыбь от лодки на поверхности бурного Атлантического океана».
Большим достижением на пути изучения пигмента — гемоглобина после получения Фишером синтетического гематина является синтетический гемоглобин, приготовленный Хиллом из гематина и глобина.
Полученный таким образом гемоглобин не дает, однако, характерного изгиба кривой диссоциации. Мелдром и Рэштон (Meldrom, Roughton) обнаружили наличие связанного с гемоглобином фермента: фермент карбангидраза обычно присутствует совместно с гемоглобином, причем этот фермент обладает способностью ускорять реакцию между С02 и водой при образовании Н2СО, а также реакцию расщепления Н2С03 на С02 и Н20. Интересна роль меди в образовании гемоглобина, особенно в связи с эффектом, который получается при лечении препаратами меди. Из крови, по имеющимся данным, ни разу не был получен гемоглобин, свободный от меди. Эльвегжем (Elvehjem) считает, что медь обладает свойствами стимулировать образование некоторых гематиновых соединений. Женское молоко вдвое богаче медью, чем молоко коровы или козы. Медь откладывается в печени наряду с гемосидерином (железосодержащий пигмент). Последний, по мнению Кука (Соок), не является определенной химической единицей; возможно, что зерна гемосидерина несут в себе железосодержащий пигмент примерно таким же образом, как эритроциты несут в себе гемоглобин. В печени зародыша много железа, которое постепенно используется после рождения.
Основным свойством живой системы является ее раздражимость факторами внешней среды, с одной стороны, и эндогенными факторами в связи с обменом веществ — с другой. Риккер утверждает, что в многоклеточном организме функциональное раздражение идет всегда через сосудисто-нервный аппарат.
Наши наблюдения убеждают нас в правильности этой теории. Вряд ли можно в каком-либо процессе установить непосредственное избирательное «специфическое» действие определенного агента на клетки органов и тканей. Действие любого раздражителя — будет ли он физиологическим или патологическим — передается клеткам, трансформированным через посредство соответственных нервных волокон. Последние в процессе филогенеза сенсибилизируются и перенимают на себя рецепторную функцию, тогда как клеточные элементы в процессе филогенеза, наоборот, теряют способность специфически реагировать на непосредствен но действующие на них раздражения, оставаясь сенсибилизированными только к нервным импульсам.
Местные процессы реактивного характера осуществляются, нужно думать, путем короткой дуги аксонного рефлекса, а не через центральную нервную систему. Раздражение нервно-сосудистого аппарата сообщается окружающей ткани; изменяясь под влиянием этого раздражения, ткани начинают или усиленнее, или качественно иначе питаться, т. е. нервно-сосудистый аппарат управляет тканевым обменом. Невроваскулярная теория клеточного раздражения получила подтверждение в недавно вышедших работах из лаборатории Штёра (Stohr); авторам удалось впервые с помощью усовершенствованной техники обработки нервных волокон выявить богатую неврогенную сеть, пронизывающую эндотелий капилляров.
Проверяя на практике теорию Риккера, мы должны были внести в нее существенные коррективы. По мнению Риккера, эндотелий капилляров играет роль исполнительного органа, отвечающего на раздражение только посредством заложенного в нем аппарата нервных окончаний; Риккер отрицает за клетками многоклеточного организма первичную раздражимость, он называет ее «метафизиологической». В этом заключается основное различие между теорией Риккера и тем, как мы понимаем ответную реакцию организма на раздражение. Наши наблюдения, основанные на изучении патологоанатомического материала (трупов людей и экспериментальных животных), а также на данных, полученных при культивировании тканей in vitro, убеждают нас в том, что паренхима специфически дифференцированных органов в процессе филогенеза тем более изолирована от внешнего воздействия, чем большее значение имеет этот орган для организма. Максимально совершенной системой изоляции от непосредственного воздействия как экзогенных, так и эндогенных факторов снабжены: 1) клетки половых желез, 2) клетки центральной нервной системы. Наименее изолированной от непосредственного воздействия раздражителя является ретикуло-эндотелиальная система. В противоположность Риккеру мы считаем, что в многоклеточном организме сохранилась первичная раздражимость клеточной протоплазмы, но это свойство перешло всецело и строго дифференцировалось в системе терминального обмена веществ органов и тканей, в эндотелии соответственных капилляров.
Нервные волокна и их окончания нигде в организме не бывают голыми — тончайшие их разветвления проходят в шванновском синцитии, сопровождающем их вплоть до клеток, которые они иннервируют и в которых они оканчиваются. Роль шванновской оболочки в эндотелии капилляра играет его протоплазма. Эндотелий капилляров является рецептором и трансформатором раздражений, получаемых им из протекающей крови, с одной стороны, и от метаболитов специфической паренхимы — с другой. Под влиянием раздражений в протоплазме эндотелия капилляра образуются различные специфические раздражители для пронизывающих протоплазму нервных волоконцев; таким образом, осуществляется расширение и сужение капилляров. Вещества, действующие на эндотелий капилляров, оказывают на него, кроме того, свое специфическое действие; например, под влиянием наркотиков капилляр приходит в состояние покоя, который характеризуется сужением просвета и пониженной проницаемостью эндотелиального барьера; функционально это выражается в резком снижении деятельности соответственного органа, например, кора головного мозга засыпает. Избирательное действие различных веществ как эндогенных, так и экзогенных на различные капилляры объясняется тем, что каждый орган, каждая ткань имеет свои специфические капилляры.
На специфичности капилляров основаны чрезвычайно ценные данные Л. С. Штерн о гемато-энцефалическом барьере и о других гисто-семитических барьерах. «Капилляры, — говорит Штерн, — не могут рассматриваться как нечто единое, идентичное по своей структуре, по своим физиологическим особенностям во всем организме. Каждая ткань имеет свои особые капилляры, которые отличаются от капилляров других органов своей морфологической структурой и биологическими особенностями».
Таким образом, мы видим в капилляро-тканевом нейро-эндотелиалъном барьере субстрат, реагирующий на эндо-экзогенные раздражители и отвечающий на него, нужно думать, согласно закону ступеней Риккера:
1. Слабое раздражение путем возбуждения сосудорасширителей ведет к расширению капилляров и к ускорению тока крови.
2. Средней силы раздражение путем возбуждения сосудосуживающих нервов ведет к сокращению капилляров и артерий и к замедлению скорости тока в капиллярах и в венах.
3. Более сильное раздражение ведет к закрытию просвета мелких артерий и капилляров. В венах ток крови останавливается.
4. При очень сильном раздражении наступает резкое расширение капилляров, ввиду потери возбудимости констрикторов, в то время как большие артерии сокращаются. Ток крови замедляется, наступает пре-статическое состояние, красный стаз, а затем и белый стаз с выхождением лейкоцитов и плазмы крови.
Встреча токсина с клетками организма происходит в эндотелии капилляров [Дёрр (Doerr)]: тут, нужно думать, происходит и обезвреживание токсина. Таким образом, действие токсина на органы происходит в основном без непосредственного повреждения специфической паренхимы и поэтому оно носит преимущественно функциональный характер. В этом мы видим причину не повреждаемости зародышевой клетки и залог прогрессивного развития центральной нервной системы. Только после повреждения или гибели нейро-эндотелиального капилляро-тканевого барьера пораженного органа токсин получает непосредственный доступ к клеткам паренхимы. Капилляро-тканевой барьер в ретикуло-эндотелиалъной системе является или частью ее функциональной единицы (как прибрежные клетки, купферовские клетки, эндотелий венозных синусов кроветворных органов), или же — в остальной части ретикулярной ткани — взаимоотношения имеются те же, что и в других тканях, но проницаемость барьера здесь совершенно иная в связи с функцией обезвреживания и накопления ретикулярной ткани.
Таким образом, болезнь заключается в изменении нормального состояния организма благодаря перестройке его различно функционирующих систем. Эта перестройка носит чисто защитный характер и при благоприятном течении любого заболевания может ограничиваться соответственными функциональными изменениями. Мы видим здесь аналогию с токсикозом, возникающим в процессе нормальной, физиологической, жизнедеятельности организма — утомление приводит функционирующие системы в состояние физиологического наркоза, сна, во время которого уничтожаются токсины утомления. В зависимости от характера и силы действующего токсина, при возникшем заболевании могут образоваться на различных участках очаги воспаления, иногда некроза. Эта очаговая воспалительная реакция приобретает особое значение лишь в некоторых случаях, в зависимости от локализации воспалительных очагов (мозг, сердце и др.) и от течения воспалительного процесса, когда воспалительный очаг становится источником нового заболевания.
Если мы рассмотрим с этой точки зрения различные инфекционные заболевания, то увидим, что так называемые локальные воспалительные изменения соответственно месту проникновения в организм токсического начала не являются первичными. Так, например, ангина при скарлатине или дифтерии развивается через известный инкубационный период после вхождения патогенного начала в организм и является уже ответом сенсибилизированных тканей на общее заболевание организма. Повреждение кожи на месте укуса клеща превращается в воспалительный фокус одновременно с развитием общего заболевания — так называемого клещевого сыпного тифа (Давыдовский).
Технические трудности при изучении вегетативной иннервации различных паренхиматозных органов чрезвычайно велики; это видно из того, что каждому из наших лучших нейрогистологов, так же как и зарубежным нейрогистологам, удается добиться ясных, четких препаратов только в каких-либо органах или тканях; на основании работ Догеля, Лаврентьева, Снесарева, Зазыбина, Леонтовича, Серебрякова, Штёра, Бука (Воеке) и др., в настоящее время все органы и ткани оказались иннервируемыми.
Относительно иннервации костного мозга в последнее время имеется значительное количество работ [Мисколши, Любош, Фоа (Miskolczy, Lubosch, Foa)]; обнаруживаемые гистологически нервы не являются только вазомоторами — нежные нервные волоконца разветвляются в паренхиме костного мозга [де Кастро, Росси (de Castro, Rossi)], что и является морфологическим выражением влияния нервных импульсов на функцию костного мозга (см. схему, рис.).
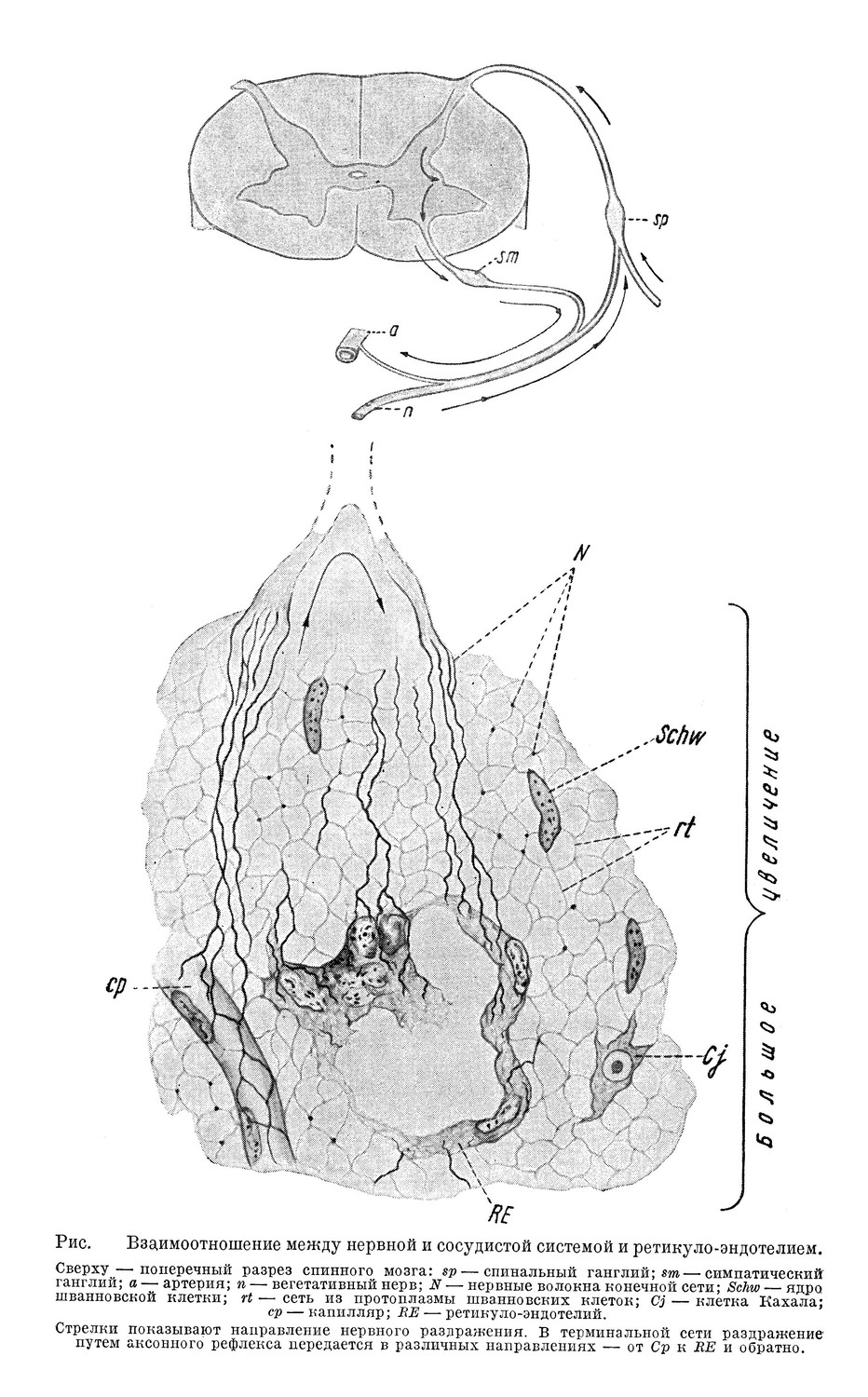
Получены также тончайшие разветвления нервных волокон в протоплазме купферовских клеток, в ретикулярном синцитии пульпы селезенки; таким образом, подведена точная анатомическая база для представления о взаимоотношении между ретикуло-эндотелиальной и нервной системой.
В настоящее время много пишут о том, что кроветворение имеет свои специальные центры, заложенные в субталамической области. Изучая гистологически субталамическую область (в которой заложен ряд вегетативных ганглиев) при различных заболеваниях крови, например, при пернициозной анемии, полицитемии, лейкемии, различные авторы находили изменения, особенно в ганглиозных клетках ядер этой области, и считали найденные изменения причиной, патогенетически обусловливающей то или иное заболевание крови. Мы в течение ряда лет систематически изучали дно третьего желудочка мозга (субталамическую область) при так называемых болезнях крови и при других заболеваниях. При этом мы убедились в том, что сосуды субталамической области чувствительны к целому ряду токсических воздействий (по типу гетеро-сенсибилизации, обусловленной физиологической, функциональной сенсибилизацией); нарушение кровообращения — стазы, кровоизлияния — имеют здесь свою излюбленную локализацию. Что касается ганглиозных клеток вегетативных ядер субталамической области, то в nucleus paraventricularis и nucleus supraopticus, как правило, мы находили большее или меньшее количество включений, главным образом в виде вакуолей; в некоторых случаях эти «изменения» были так резко выражены, что импонировали как серьезное поражение. Работы последних лет Петерса, Русси (Peters, Roussy) и др. убеждают нас в том, что в ганглиозных клетках вегетативных ядер вакуоли и включения являются продуктами нормальной внутренней секреции этих клеток (нейрокриния), быть может, связанной с гормоном гипофиза. Особенное внимание взаимоотношению между кроветворением и дном третьего желудочка мозга стали уделять после того, как после энцефалографии в ряде случаев были обнаружены лейкоцитозы, сопровождавшиеся другими признаками вегетативного характера (например, гипертермией). Экспериментом на животных [Порта, Греко (Porta, Greco)] пытаются доказать, что раздражение паравентрикулярного ядра ведет к лейкоцитозу, лимфопении и анемии, тогда как разрушение его сопровождается полиглобулией и лейкопенией. Изменения со стороны лейкоцитов при сотрясении мозга, при шоке и, наконец, тромбопению с явлениями геморрагического диатеза также связывают с поражением гипоталамо-гипофизарной системы. Лихвиц (Lichwitz) высказывает даже предположение, что агранулоцитоз, развивающийся в некоторых случаях после употребления пирамидона, также гипоталамического происхождения.
На основании имеющихся в настоящее время гистологических данных и данных эксперимента функциональная связь между капилляром, паренхиматозными клетками и кровеносными сосудами осуществляется следующим образом: по Штёру, единство капилляра и паренхиматозной клетки обусловливается терминальным сплетением; по Бейлису (Beyliss), в чувствительных волокнах, проходящих через задний корешок спинного мозга, имеются волокна, которые, отщепляясь, присоединяются к симпатическому нерву и, ветвясь, оканчиваются частью в клетке паренхиматозного органа, частью в питающем их капилляре. Чувствительное раздражение, идущее центрипеталъно у места разветвления, принимает обратное направление и антидромно достигает капилляра, функционируя как вазодилятатор, т. е. путем аксонного рефлекса осуществляется функциональное единство капилляра и паренхиматозной клетки. Аналогичную картину иннервации Лаврентьев наблюдал в гладкой мускулатуре кишечника.
Согласно взглядам Сперанского, нервной трофикой может обладать всякий нерв. Вся нервная система по его учению представляет одно неразрывное целое и выключение деятельности одного участка перестраивает деятельность всей нервной системы таким образом, что она иначе реагирует на раздражение. Локализация дистрофии не играет, по его мнению, никакой роли.
На схеме (рис.) представлены взаимоотношения между нервной и сосудистой системой и клетками паренхимы. Кровеносный капилляр и прилегающая к нему петля ретикулярной ткани (RE) кроветворного органа соединены между собой окутывающей их сетью протоплазматического синцития, в котором заложены нервные волоконца — так называемая концевая, или терминальная, сеть; в последней заложены [по Кахалу (Gajal)] «интерстициальные нервные клетки», трансформирующие раздражение и, возможно, вырабатывающие специфические вещества. Как видно из схемы, иннервация капилляра, согласно работам Бейлиса, осуществляется волокнами, которые симпатический нерв получает из межпозвоночного (спинального) ганглия. Таким образом, осуществляется координация функции паренхиматозных клеток и их капилляров. Протоплазма эндотелия капилляра воспринимает раздражение, приносимое ей током крови, и передает его нервным окончаниям. Продукты утомления, например, как было указано выше, выявляют свое действие на эндотелии капилляров и таким образом снижается функция паренхиматозных клеток — засыпают, согласно такому пониманию, эндотелиальные клетки капилляров мозга, а ганглиозные клетки благодаря этому приходят в неактивное состояние, состояние физиологического сна. Тормозное действие какого-либо вещества на эндотелий капилляров костного мозга выявляется прекращением или понижением функции его ретикуло-эндотелия.
Эта краткая экскурсия в область общебиологических вопросов облегчит нам выяснение тех задач, которые встают перед нами при изучении морфологии клетки вообще и кровяных телец в частности.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
|
