Уже И. И. Мечников при сравнительно патологическом изучении воспаления совершенно отчетливо выделил и описал клеточные элементы, принимающие участие в фагоцитной реакции организма. В первую очередь им подчеркнута сократимость «эндотелиальных» клеток, которые могут оставлять сосудистую стенку и с помощью амебоидных движений проникать в полость сосудов. Это особенно часто наблюдается в капиллярах печени: эндотелиальные клетки печеночных капилляров легко отрываются от адвентиции и появляются в форме звездчатых клеток. Они легко захватывают некоторые виды микробов и другие «посторонние тела». Теми же свойствами обладает и эндотелий лимфатических узлов (Риберт — Ribbert).
Помимо этого, И. И. Мечников считал, что одноядерные лейкоциты (судя по иллюстрации — моноциты) несомненно обладают фагоцитарными свойствами и «становятся эпителиоидными, грануляционными и гигантскими клетками».
При изучении туберкулеза как хронического воспаления он показал, что в образовании туберкула в печени «никогда ни одна печеночная или эпителиальная клетка не способствует образованию туберкула.» Эпителиоидные и гигантские клетки образуются только за счет одноядерных лейкоцитов и звездчатых клеток Купфера. Внутри этих фагоцитов находятся бациллы. Сливаясь, эпителиоидные клетки образуют гигантские клетки. Сегментноядерные лейкоциты, фагоцитируя туберкулезные бациллы, гибнут или же сразу фагоцитируются различными одноядерными фагоцитами — макрофагами. При введении гусиных эритроцитов в брюшную полость морской свинки появляется большое количество макрофагов, поглощающих эти эритроциты. Вместе с тем было отмечено, что участие клеток соединительной ткани менее важно и менее активно в сравнении с деятельностью лейкоцитов и эндотелиальных клеток кровеносных сосудов.
Роль неподвижных клеток соединительной ткани сводится к замещению элементов ткани, разрушенных при воспалении. Таким образом, уже И. И. Мечников выделил различные элементы, обладающие фагоцитными свойствами, поглощающие не только бациллы, но и клеточные элементы и их распад, и объединил их по этому свойству общим именем «макрофагов».
В то же время (1890—1896 гг.) Ранвье (Ranvier) описал неподвижные клетки, обнаруженные им в рыхлой соединительной ткани, способные выпускать отростки и названные поэтому клазматоцитами. При раздражении они размножаются и становятся подвижными. Им приписывали функции фагоцитов, переносчиков питательных веществ, и полагали, что они являются производными лейкоцитов, мигрировавших из сосудов в ткани. Способность этих клеток к интенсивному фагоцитозу подчеркивалась особенно В. К. Высоковичем и И. И. Мечниковым, включавшими их в число макрофагов. Клетки, подобные клазматоцитам, были описаны Маршаном как адвентициальные и А. А. Максимовым как поли бласты.
Особое значение для изучения и оценки этих клеточных элементов имели работы Гольдмана (Goldmann). На основании своих исследований с введением пирроловой и трипановой сини, отлагавшихся в ретикулярных и эндотелиальных клетках различных органов он пришел к заключению, что, с одной стороны, эти элементы обладают общностью функций в процессе обмена веществ, и, с другой, установил идентичность своих пирроловых клеток с клазматоцитами Ранвье, адвентициальными клетками Маршана, полибластами Максимова и т. д.
Эти исследования послужили основанием для огромного количества работ, посвященных изучению с различных точек зрения способности некоторых клеток к прижизненному накапливанию коллоидов, липоидов и пигментов. Эти работы привели к выделению распространенных в организме млекопитающих определенных видов клеток, обладающих указанной общностью функций, и позволили сгруппировать их в одну систему, которую назвали «эндотелиальным обменным аппаратом» (Ашофф-Ландау — Landau), гистиоцитарным обменным аппаратом (Кионо — Kijono). В дальнейшем и по настоящее время за этим аппаратом сохранилось название — ретикулоэндотелиальная система.
Клетки этой системы, как указывает Бернер-Патцель (Boerner-Patzelt) обладают такими общими свойствами:
1) способностью играть роль при кровообразовании в эмбриональной и последующей жизни, в особенности при патологических условиях;
2) способностью к интенсивному фагоцитозу;
3) способностью накапливать в себе большое число электроотрицательных коллоидных веществ, которые вводятся в организм хотя бы в небольшом количестве и слабой концентрации.
Особо нужно подчеркнуть способность этих клеток прижизненно поглощать красящие вещества, причем последние откладываются в виде равномерного зернистого осадка без какого- либо повреждения клетки.
Из сказанного достаточно ясно вытекает определение понятия ретикулоэндотелиальной системы, которое сформулировано Станденатом (Standenath) следующим образом: с физиологической и патологической точки зрения под названием ретикулоэндотелиальной системы мы понимаем рассеянные по всему телу, но сосредоточенные в определенных местах скопления мезенхимальной ткани, которые в биологическом отношении характеризуются способностью к внутриклеточному накапливанию самых различных электроотрицательных коллоидов неорганической и органической природы, экзогенного и эндогенного происхождения.
В систему ретикулоэндотелия включены мезенхимальные элементы, которые по степени прижизненной окраски Ашофф подразделяет на четыре группы (из шести мезенхимальных элементов, способных вообще к поглощению).
1. Эндотелий кровяных и лимфатических сосудов; он поглощает только при особо интенсивной окраске и только в виде мельчайших зернышек.
2. Фиброциты, или обычные соединительнотканные клетки; они поглощают при достаточно сильной окраске с различной силой, но также достаточно мелкозернисто, однако легче красятся, чем эндотелий.
3. Ретикулярные клетки селезеночной пульпы, фолликулов, коры и мозговых тяжей лимфатических узлов и прочей лимфатической ткани. Они поглощают относительно легко и сильнее, чем клетки соединительной ткани, но еще явно отстают по быстроте и силе накопления от следующих групп.
4. Ретикулоэндотелий лимфатических синусов, лимфатических узлов, кровяного синуса селезенки, капилляров печеночных долек (купферовские звездчатые клетки), каппиляров костного мозга, коры надпочечника, гипофиза.
5. Гистиоциты, как мы обозначаем подвижные элементы соединительной ткани, клазматоциты Ранвье и т. д. В противоположность другим образованиям соединительной ткани (фибробластам, фиброцитам) они поглощают почти также легко, как клетки четвертой группы, особенно когда находятся в деятельном состоянии.
6. Спленоциты и поглощающие красящие вещества моноциты (эндотелиолейкоциты, кровяные гистиоциты), которые происходят из гистиоцитов (группа 5) и ретикулоэндотелия (группа 4).
В тканях они расположены изолированно или небольшими группами, представляя клетки неодинаковой величины и формы — овальные, круглые, вытянутые в одном направлении, или отростчатые, часто с включениями в протоплазме. Еще более разнообразны гистиоциты в очагах воспаления, где они встречаются то в виде очень маленьких клеток, с трудом или вовсе не отличных от малых лимфоцитов, то в виде очень крупных клеток, содержащих многочисленные включения. Ядро гистиоцитов относительно мало, окрашивается более интенсивно, чем ядра других соединительных клеток, овальной или почковидной формы и обладает толстой, складчатой мембраной (Зильберберг — Silberberg). В ядре определяются грубые, неравномерно расположенные частицы хроматина и часто неясно контурирующееся ядрышко. Протоплазма темная, имеет грубоватое зернисто-сетчатое строение и содержит часто разнообразные включения, вакуоли, жировые капли. Около ядра ясно вырисовывается клеточный центр.
По способности интенсивно накапливать коллоидные краски и по морфологическим свойствам гистоциты хорошо отличаются от других тканевых элементов. По сути это те клетки, которые находятся в кроветворных органах и описаны ниже среди ретикулоэндотелиальных элементов как фагоцитирующие ретикулярные клетки — макрофаги и лимфоидные ретикулярные клетки. Гистиоцитам принадлежит большая роль в развитии, течении и исходе воспаления, морфологические изменения при котором очень хорошо прослежены и систематически описаны А. Максимовым, М. Зильбербергом (Silbezberg) с приложением прекрасных иллюстраций, и в 1948 г. В. Менкиным. И. И. Мечников, А. А. Максимов, М. Зильберберг, В. Менкин и другие авторы с полной очевидностью показали, что в первые часы, наряду с предсуществующими тканевыми элементами, появляются и нарастают в количестве мигрирующие через стенку сосудов полиморфноядерные лейкоциты, отграничивающие очаг воспаления вокруг инородного тела.
Наряду с полиморфноядерными лейкоцитами через стенку сосудов мигрируют и лимфоциты. В работе и иллюстрациях М. Зильберберга подчеркивается, что лимфоциты скоро меняют свое существо. Если в первый момент они сохраняют еще вид типичных малых лимфоцитов, то в последующие 6 — 8 часов протоплазма их гипертрофируется, а через 12—24 часа лимфоциты изменяются еще больше. Оболочка их ядра становится тоньше, хроматиновые частички мельче, протоплазма шире. Большая часть лимфоцитов претерпевает указанные превращения в полибласты, а меньшая часть дегенерирует. В это время появляется все больше гистиоцитов макрофагов, которые примешиваются к мигрировавшим из крови лейкоцитам. Таким образом, в воспалительном очаге можно различить двоякого рода одноядерные элементы. Но уже через 36 часов лимфоциты принимают различную, не совсем правильную форму, хроматиновая сетка ядра становится более рыхлой и в противоплазме их обнаруживаются различные включения — остатки лейкоцитов, эритроцитов и т. п. По данным А. Максимова, М. Зильберберга и других авторов трудно отличить образовавшиеся из лимфоцитов гематогенные полибласты, превращающиеся в макрофагов, от гистиогенных блуждающих клеток. В дальнейшем и те и другие превращаются в очаге воспаления в фибробласты.
Перечисленные выше авторы, как и другие гистологи и патогистологи, единодушно приписывают лимфоцитам значительную роль в образовании макрофагов. Вместе с тем это положение не кажется нам достаточно ясным. С того времени, как А. Максимов заменил определение материнской клетки как «малый лимфоцит» на лимфоидный «гемицитобласт», большинство гематологов и многие гистологи считают малый лимфоцит вполне дифференцированной кровяной клеткой, неспособной к превращениям. С этой точки зрения образование полибластов — гистиоцитов из лимфоцита мало вероятно. Кроме того, И. И. Мечников и другие исследователи, подчеркивают, что из лейкоцитов наименьшей способностью образовывать протоплазматические отростки и подвижностью обладают лимфоциты. Последние не являются фагоцитами. К тому же одним из условий, благоприятствующих миграции лейкоцитов, является сегментация ядра, что не наблюдается у лимфоцитов, обладающих сравнительно крупным, компактным ядром. Эти обстоятельства вместе с тщательным изучением работ А. Максимова, Ланга, Блюма, А. Д. Тимофеевского и С. В. Беневоленской и других авторов внушают некоторое сомнение в правильности трактовки лимфоидных элементов как малых лимфоцитов. Для иллюстрации мы приводим цветной рисунок А. Максимова, где показан последовательный ряд превращений малого лимфоцита в амебоидный полибласт, накапливающий кармин. Ниже приведено описание морфологических особенностей так называемых лимфоидно-ретикулярных клеток, которые в количестве от 1 до 3% могут обнаруживаться в крови даже при физиологических условиях. Изображенный на рисунке А. Максимова лимфоцит полностью соответствует представлению о лимфоидно-ретикулярной клетке, и тогда не удивительно, что она превращется в полибласт и фибробласт.
В клеточных реакциях при нормергическом воспалении, как это классически было описано И. И. Мечниковым, основное количественное и функциональное значение принадлежит сегментоядерным нейтрофилам—микрофагам, но наряду с ними принимают определенное участие в воспалительном процессе указанные гистиоцитарные элементы — макрофаги. Несколько иная картина наблюдается при гиперергической реакции.
М. А. Скворцов подчеркивает правильность характеристики гиперергической воспалительной реакции, данной Герляхом (Gerlach), который считает, что принципиальной разницы между нормэргической и гиперергической реакцией не существует, и суть заключается только в изменении степени (т. е. повышении интенсивности всех явлений и быстроте). Исходя из этого положения, М. А. Скворцов подчеркивает, что при гиперэргических воспалительных реакциях, как и при обычных, можно ясно наметить три категории изменений:
1) группу с преимущественно альтеративными (дегенеративно-некротическими изменениями),
2) группу с преимущественно экссудативными изменениями (отек, фибрин, нагноение)
3) группу с изменениями, главным образом, пролиферативного характера.
Принадлежность к той или иной группе зависит от интенсивности реакции. Существенную роль в развертывающейся картине поражения играет нарушение функции сосудистой системы. Обширный стаз, кровоизлияния, тромбоз будут способствовать некрозу; при меньшей силе вредного воздействия на сосуды будет преобладать образование .экссудата, жидкого или клеточного; при еще более слабом поражении сосудов и меньшем повреждении окружающей ткани на первый план выступят явления клеточного размножения.
В клинической патологии наблюдаются преимущественно вторая и третья группы. При этом в третьей группе с явлениями, главным образом, продуктивными, характерными являются не столько количественные изменения, сколько своеобразное образование узелка — гранулемы.
Для изучения таких изменений при типе воспаления, которое определялось как хроническое, И. И. Мечников избрал в качестве примера, как уже было указано, образование туберкулов при чахотке. И. И. Мечников приходит к выводу, что «туберкул состоит из скопления фагоцитов мезодермического происхождения; эти фагоциты приходят к месту нахождения бацилл и захватывают их. Фагоциты остаются в форме эпителиоидных или превращаются в гигантские клетки. Последние могут развиваться различными способами, приводящими всегда к образованию больших протоплазматических масс, заключающих многочисленные ядра». По существу к сказанному мало что добавлено дальнейшими исследованиями. Резюмируя ряд работ по гистологии туберкула, Н. Н. Аничков пишет: «Из приведенных до сих пор работ видно, что участие клеточных элементов ретикулоэндотелиальной системы — гистиоцитов в образовании бугорка выражается в превращении их в эпителиоидные и затем частью в гигантские клетки.»
Нет нужды останавливаться на изменениях при других типах гранулом, которые отличаются друг от друга только в деталях. Таким образом, всякая гранулома является ретикулоэндотелиальным образованием, при чем ретикулоэндотелиальные клетки превращаются в эпителиоидные и часто в гигантские клетки. В настоящее время уже не остается сомнений в том, что гранулома является одним из проявлений гиперергической воспалительной реакции с изменениями, главным образом, пролиферативного характера.
Клетки ретикулоэндотелиального аппарата могут отделяться и поступать в кровь, причем часть из них проходит короткий путь до легких, где они задерживаются и разрушаются. Другая часть поступает в легочные вены и разрушается в сердце. Таким образом, только очень небольшое количество сохранившихся ретикулоэндотелиальных элементов поступает в общий круг кровообращения и циркулирует в нем.
Эти свободные гистиоцитарные клетки причисляются обычно к кровяным моноцитам или отождествляются с ними. Вместе с кровяными моноцитами эти клетки расцениваются как кровяные элементы, в связи с чем в последнее время и возникло учение Шиллинга о третьем виде клеток крови — моноцитах, как производных отдельной кроветворной ткани, которую можно противопоставить миэлоидной и лимфоидной.
Шиллинг считает, что ретикулоэндотелиальная система является не только функционально объединенной тканью, но одновременно и третьей самостоятельной кроветворной тканью, производящей клетки третьей группы лейкоцитов — моноциты.
А. Максимов и его школа объединяют моноциты в одну группу с большими родоначальными лимфоцитами. Своими исследованиями Блюм доказывает, что, с одной стороны, in vivo и особенно в тканевых культурах моноцитоподобные клетки развиваются из лимфоцитов нормальной и лейкемической крови, а также из миэлобластов лейкемической крови. С другой стороны, моноциты очень быстро превращаются в тканевых культурах в макрофаги — фибробласты. Особенно доказательны в этом отношении исследования А. Д. Тимофеевского и С. В. Беневоленской. При отсутствии митозов моноцитов они находили большое количество моноцитоподобных клеток, которые являлись переходными формами от лимфоцитов к полибластам. Паппенгейм и Феррата (Ferrata), на основании исследования крови кроликов, продемонстрированного прекрасными таблицами, приходят к выводу, что большие лимфоидные лейкоциты (моноциты) происходят из больших нормальных лимфоцитов вследствие простого их старения. Если для нормальной крови эти авторы принимают происхождение моноцитов только от нормальных больших лимфоцитов, то для патологической крови допускают существование, так называемых, патологических моноцитов, под которыми подразумевают стареющие лейкобласты, не считая их законченными формами, не могущими дифференцироваться дальше, в противоположность нормальным лимфоидным моноцитам. Гейнц (Heinz) и Циглер (Ziegler) подобно Паппенгейму, считали моноциты стареющими формами миэлобластов, способными все же к нормальной дифференциации.
Ортодоксальные дуалисты (Негели, Тюрк, Альдер и др.) считают, что моноцит это особая клеточная форма, происходящая от миэлобласта и близко к нему стоящая, но отличающаяся от него морфологически и биологически. Шиллинг, Негели, Гиттмайер находят, это развитие от миэлобласта к кровяному моноциту проходит через стадию монобласта, т. е. клетки с более базофильной, чем у зрелого моноцита протоплазмой и юным, содержащим нуклеоли, ядром.
Особое мнение относительно происхождения моноцитов, которое не находило в свое время призерженцев, было высказано Пателла (Patella). По его теории, моноциты происходят не из гематопоэтических органов, а являются десквамированным эндотелием сосудов.
Вопрос о моноцитах получил освещение со времени опубликования работ Ашофф и Кионо. На основании своих исследований с инъекциями литиокармина они приходят к заключению, что нужно различать три вида лейкоцитов:
1) миэлолейкоциты (миэлобласта, миэлоциты),
2) лимфолейкоциты (лимфоциты)
3) гистио- или эндотелиолейкоциты (клазматоциты Ранвье и адвентициальные клетки Маршана).
Эти исследователи, вместе с Ванденрайхом, считают, что клазматоциты (гистиоциты) функционально равнозначущи клеткам эндотелия лимфатических синусов и капилляров селезенки, печени и костного мозга. Кионо в своих опытах отметил факт увеличения в крови количества мононуклеаров и переходных форм, поглотивших краску (гистиоцитемию). При ближайшем изучении этих клеток оказалось, что ни по форме, ни по структуре ядра и протоплазмы отличие этих клеток от обычных моноцитов было бы новозможно без витальной окраски. Моноциты (мононуклеары и переходные формы), которые составляют у кролика 5—10% общего количества лейкоцитов в крови периферических сосудов, в большинстве не представляют собой гистиоцитов. После инъекции краски гистиоциты в небольшом количестве проникают в кровь, моноциты же крови красок не поглощают, но, как и лимфоциты, могут нарастать в процентом отношении.
Разбирая на основании своих исследований вопрос о происхождении моноцитов и их отношении к гистиоцитав, Кионо приходит к заключению, что мононуклеарные клетки гематопоэтических органов по их поведению при инъекциях толуидинблау разделены на три вида: прижизненно зернистые гистиоциты, поствитально гранулированные миэлоидные клетки и при жизни и поствитально беззернистые клетки, соответствующие лимфоцитам и лимфобластам. Соответственно этим трем группам существуют, по крайней мере, три функционально и генетически различных вида моноцитов. Наши наблюдения согласуются с данными Кионо, и мы целиком поддерживаем его выводы.
Как ни различны, таким образом, отдельные взгляды на происхождение моноцитов, они имеют то общее, что не признают моноцит кровяной клеткой из отдельной кроветворной системы, наряду с миэлоидной и лимфатической.
Из сказанного можно сделать вывод, что моноцит не представляет собой отдельной клеточной формы и является понятием собирательным для клеток внешне подобных, но разного происхождения, и не всякую клетку, попадающуюся в циркулирующую кровь, можно считать кровяным элементом.
Данные современных клинико-морфологических методов прижизненного исследования кроветворных органов с помощью пункции костного мозга, лимфатических узлов, селезенки и печени не дают никаких указаний на происхождение моноцитов из ретикулярной ткани, основная масса которой в организме сосредоточена в кроветворных органах. Прижизненное исследование кроветворных органов не дает указаний также на непосредственное участие ретикулума в гемопоэзе при нормальных условиях. Каждая гемопоэтическая система — лейкоцитарная, эритроцитарная, тромбоцитарная имеет в кроветворных органах свою родоначальную функционально более или менее направленную клетку — гемоцитобласт, проэритробласт, мегакариобласт, близкие по строению ядра и базофилии протоплазмы. Количество таких родоначальных элементов в кроветворных органах невелико по сравнению с более зрелыми, но способными еще к размножению, более строго дифференцированными элементами — эритробластами, лейкобластами, миэлоцитами, лимфобластами, мегака- риоцитами, обеспечивающими нормальный состав крови. При физиологических условиях ретикулярная ткань является только опорной тканью, связанные или свободные элементы которой обладают чрезвычайно важными для организма функциями, о которых речь будет идти ниже. При повышенных требованиях ретикулярная ткань отвечает не только повышением функции, но и гиперплазией. Только при особых, не совсем еще ясных влияниях, ответная реакция элементов ретилулярной ткани проявляется в их эмбриональной полипотентной способности образования, в частности, кровяных элементов, характеризующих патологическое состояние кроветворной системы.
Большинство современных исследователей принимает предложенное Рором подразделение ретикулярных элементов в нормальном костном мозгу на четыре основные группы:
1) фагоцитирующие ретикулярные клетки (макрофаги),
2) лимфоидные ретикулярные клетки,
3) плазматические ретикулярные клетки,
4) юные ретикулярные клетки.
Мы совершенно не разделяем воззрений Рора, который смотрит на плазматические клетки только как на функционально различную форму ретикулума, а рассматриваем их как вполне дифференцированный функционально и морфологически элемент костного мозга и описываем их в соответствующем месте. Мы не можем также согласиться с положением Негели и его школы, что клетки Феррата это механически поврежденные промиэлоциты или миэлоциты. Наблюдения над различными патологическими процессами в кроветворных органах, протекающими с их гиперплазией и изучение эмбрионального гемопоэза позволяет полагать, что гемогистиобласт — форма ретикулума с определенной направленностью к превращению в кровяные элементы.
Лимфоидно-ретикулярные клетки составляют большую часть ретикулярных клеток костного мозга, но они могут встречаться часто в крови и должны быть отдифференцированы от лимфоцитов. Таким образом, мы подразделяем ретикулярные элемнты на четыре группы:
1) юные ретикулярные клетки;
2) фагоцитирующие ретикулярные клетки — макрофаги;
3) лимфоидно-ретикулярные клетки;
4) гемогистиобласты и гемогистиоциты (клетки Феррата).
Юные ретикулярные клетки
Юные ретикулярные клетки представляют собой более или менее правильной формы круглые или овальные клетки со сравнительно большим ядром и меньшим количеством протоплазмы. Ядро напоминает по структуре ядро гемоцитобласта. Оно также имеет зернистое строение, но состоит из более грубых зернышек. По аналогии с гемоцитобластами нужно полагать, что зернистая структура является выражением утолщения на местах пересечения нитей хроматина. На более поздних степенях развития структура выражается нежной, мелкой петлистостью и обычно в центре имеет продольное вдавление, которое придает ядру характерный вид (табл. 1). В ядре часто заметно одно или два синеватых ядрышка. Протоплазма синеватого цвета, значительной интенсивности в более юных и меньшей интенсивности в более зрелых элементах, зернистости не содержит. Юные ретикулярные клетки превалируют при особенно остром ретикулоэндотелиозе. Таким образом, можно полагать, что и ретикулоэндотелиальные клетки при своем образовании из элементов индифферентной мезенхимы проходят свою лимфоидную предстадию, несколько отличную от гемогистиобласта по структуре, но равнозначущую ему по степени зрелости.
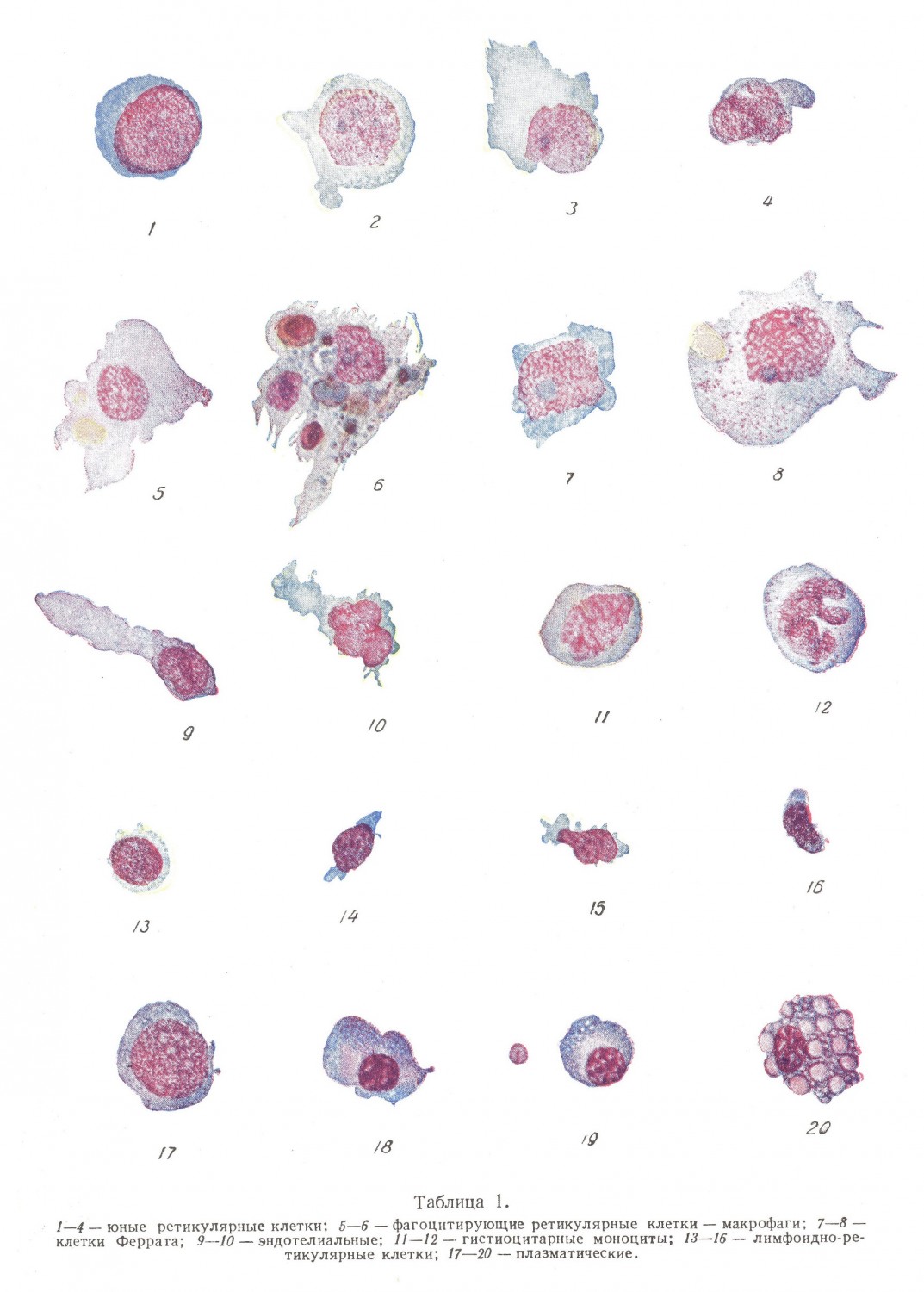
Фагоцитирующие ретикулярные клетки (макрофаги)
Фагоцитирующие ретикулярные клетки имеют различную величину и относительно малое, эксцентрично расположенное ядро. В более молодых клетках ядро имеет рыхлую, мелкопетлистую структуру, окрашивается, как и ядра миэлоидных клеток, в красно-фиолетовый цвет, с преобладанием красноватого оттенка, иногда содержит одно или два светло-синеватых ядрышка. В зрелых клетках ядро пикнотизируется. В более юных клетках протоплазма синеватая, ограниченная, в зрелых клетках она большей величины, менее очерченная, постепенно исчезающая, мало заметная и окрашивающаяся часто в розовато-синеватый цвет. В протоплазме более зрелых макрофагов, как правило, можно видеть азурофильную то более мелкую, то более крупную зернистость. В фагоцитирующих клетках в большинстве находят только глыбки пигмента грязновато-синего или бурого цвета. При повышенной функциональной деятельности в макрофагах можно обнаружить еще не потерявшие формы и окраски эритроциты и реже лейкоциты.
Лимфоидно-ретикулярные клетки
До последнего времени считали, что в периферической крови все незернистые лейкоцитарные элементы представлены лимфоцитами. Всякое изменение формы (кроме Ридеровского) ядра и протоплазмы относилось за счет механического повреждения клетки при производстве мазка. В настоящее время клинико-морфологические наблюдения с несомненностью говорят о том, что в кровь часто попадают маленькие клетки, трудно отличимые от лимфоцита, обычно с вытянутой протоплазмой и ядром, иногда более нежной структуры, чем в лимфоците, но часто мало или совсем не отличающимся от лимфоцитарного, с едва заметным ободком протоплазмы, вытянутой с одного конца или имеющей форму клина, основание которого отделено от острия круглым ядром. Клетка как бы оторвана от синцитиальной связи с другими. Протоплазма несколько более базофильна, чем в лимфоцитах и не имеет зоны просветления вокруг ядра.
Такие клетки можно обнаружить в крови в значительном количестве при разнообразных патологических состояниях, но единичные экземпляры (иногда даже до 1—2%) обнаруживаются и в нормальной крови.
Количество малых лимфоидно-ретикулярных клеток нарастает при хронических инфекциях, ретикулоэндотелиальных реакциях, ретикулоэндотелиозах, лимфогрануломатозе, инфекционном мононуклеозе.
Гемогистиобласты (клетки Феррата)
Гемогистиобласты — крупные, размером до 40 µ, круглые или полигональные клетки с круглым, овальным или неправильной формы ядром. Ядро бледное, обладает характерной структурой редкой сетки из грубых перекладин хроматина и при окраске по Гимза и Паппенгейму окрашивается в красный с фиолетовым оттенком цвет, с почти всегда ясно заметными двума-тремя ядрышками голубого цвета. Ядро эксцентрично расположено в светлой, нежноголубого цвета, широкой, неправильной формы протоплазме, которая в большинстве содержит обильную, мелкую, красную азурофильную зернистость. Своей своеобразной структурой гемогистиобласт резко выделяется среди других клеток, и его распознавание обычно не встречает затруднений. Феррата различал* по свойствам протоплазмы шесть типов указанных клеток:
1) базофильные беззернистые;
2) базофильные с азурофильной зернистостью;
3) базофильные с азурофильной и эозинофильной зернистостью;
4) утратившие базофилию с эозинофильной зернистостью;
5) базофильные с азурофильной и нейтрофильной зернистостью;
6) утратившие базофилию с нейтрофильной зернистостью.
Гиттмайр считает, что название «основные материнские клетки — гемогистиобласты» должно относиться только к первым двум типам этих клеток, не содержащим зернистости или обладающим азурофильной зернистостью; остальные четыре группы он предлагает относить к гемогистиоцитам. Феррата считает, что гемогистиобласт является основной клеткой, производящей кровяные элементы.
Мы относим к гемогистиобластам нормального костного мозга только те клетки Феррата с синеватой или голубой хорошо контурированной протоплазмой, в которых нет зернистности или содержится только скудная азурофильная зернистость. Крупные клетки в большинстве неправильной формы, с не резко очерченной протоплазмой, содержащей обильную зернистость, и грубо петлистым ядром, выделяем как гемогистиоциты. Мы считаем, что последние являются формами последующего созревания гемогистиобластов, теряющих вместе с тем свои гемопоэтические возможности. При патологических гиперпластических процессах в кроветворных органах гемогистиобласты обладают ядром более нежной структуры с крупнозернистой сеткой хроматина и более интенсивно окрашенной в синевато-голубой цвет протоплазмой. От этих клеток можно видеть постоянные переходы к гемоцитобластам.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
|
