Процесс гемолиза, т. е. выхождения гемоглобина из эритроцитов, может сопровождаться распадом, растворением основы эритроцита (stromatolysis), или же лишенные гемоглобина тела эритроцитов — тени эритроцитов — продолжают циркулировать в крови и как таковые являются патологическим началом; в последнем случае имеется только haemoglobinolysis, chromolysis. Работами Бринкмана (Brinkman) было установлено, что часть лишенных гемоглобина эритроцитов может снова заполняться гемоглобином — обратимый гемолиз (reversible haemolyse). Механизм гемолиза подвергнут детальному изучению в той мере, в какой он доступен изучению в пробирке. Однако прижизненный гемолиз в циркулирующей крови и гемолиз в пробирке во многом являются совершенно разнородными процессами, так как мы имеем в пробирке эритроциты в состоянии некробиоза; плазма крови или совсем отсутствует, или же лишена тех веществ, которые возникают в ней под влиянием введения того или иного вещества в живой организм. Поэтому и наблюдаются такие явления, что яд гемолизирует in vivo и не действует гемолитически in vitro, и наоборот. Неизвестно, происходит ли в норме более или менее значительный внеклеточный гемолиз; по-видимому, эритроциты погибают при нормальном темпе разрушения внутриклеточно, главным образом в ретикуло-эндотелии селезенки, или же они предварительно разрушаются в пульпе селезенки, где кровь застаивается.
Так как мы знаем еще чрезвычайно мало о механизме распада гемоглобина, то и методика определения количественной величины распада красной крови крайне неточна и мало убедительна. Это относится также к наиболее употребительному способу определения распада крови — к определению количества выделяемого уробилина мочой и калом, которое должно служить показателем кровяного распада.
Определяя величину распада эритроцитов по уробилину в кале, нужно считаться с трудностями самой методики и, кроме того, нужно принимать во внимание, что нет гарантии в том, что весь билирубин в кишечнике превращается в уробилин, что часть билирубина не всасывается обратно и что, наконец, билирубин не распадается на другие составные части. В толстых кишках под влиянием бактериальной флоры билирубин восстанавливается в уробилиноген и уробилин, большая часть которого выделяется калом. Билирубин в кале обнаруживается только при расстройстве моторики кишечника. В норме попадающий в печень уробилин как таковой не пропускается в общий круг кровообращения, он или разрушается или переходит в билирубин. При отсутствии усиленного гемолиза появление в моче уробилина — пигмента больной печени — служит показателем функциональной недостаточности печени. При внутривенном введении 0,05 г билирубина у здорового человека через 3 часа не должно быть гипербилирубинемии.
Вещества, обладающие гемолитическим действием, приводят к непосредственному, первичному гемолизу эритроцитов; в отличие от этого типа гемолиза, в течение целого ряда интоксикаций создаются такие условия, которые ускоряют отмирание эритроцитов, вследствие чего через различные промежутки времени наступает массовый распад эритроцитов — вторичный, или. поздний, гемолиз. Таким образом, в патогенезе позднего гемолиза отсутствует гемолитическое действие токсического вещества как такового, что и является основным отличием вторичного, позднего гемолиза от первичного гемолиза.
При патологическом гемолизе, когда в плазме крови накопляется значительное количество растворенного гемоглобина и остатки стромы эритроцитов, наступают явления интоксикации, которые различными авторами объясняются различно. Так, Гемпель (Hempel) считает, что растворенный гемоглобин действует токсически на вегетативные центры межуточного мозга, главным образом субталамической области. Гессе объясняет гемолитический шок параличом капилляров, следствием которого он считает и наступающую анурию. Возможно, что остатки эритроцитов как инородные тела вызывают озноб. Большой шаг вперед сделан в этом направлении благодаря недавно вышедшим работам Бингольда (Bingold), который доказывает, что в почках происходит значительный распад эритроцитов и гемоглобина и что пигменты мочи являются в результате распада гемоглобина. Исследования Бингольда устанавливают, что протекающая через почки кровь частично становится свободной от каталазы. Фермент каталаза предохраняет гемоглобин от действия перекиси водорода. Каталаза является образованием из класса гематинов, она превращает перекись водорода в воду и кислород (одна молекула каталазы при 40° может разрушать около 550000 молекул перекиси в 1 секунду). Перекись водорода является специфически действующим веществом, разрушающим гемоглобин.
Одним из продуктов распада гемоглобина под влиянием перекиси водорода является вещество, переходящее затем в так называемый пентдиопент (pentdyopent); это вещество обнаруживается в моче и дает спектроскопически характерную линию поглощения между 535 uu и 525 u.
Бингольд считает, что в почках имеются такие количества перекиси водорода, которые являются достаточными, чтобы производить соответственные расщепления гемоглобина. При этом гемоглобин, окисляясь, переходит в бесцветное соединение (лейкосоединение). Большая часть железа остается в циркулирующей крови, часть пигмента выделяется мочой в виде мочевых пигментов. Эти исследования Бингольда несомненно начинают новую главу о пигментном обмене. Начинается она очень важным положением, что конечными продуктами распада являются не только гематин, порфирин, билирубин и уробилин, как это считалось прежде. Ретикуло-эндотелий печени участвует преимущественно в перерабатывании шлаков эритроцитов. Резко изменяется процесс, когда в организм попадают гемолитические яды.
Имеющиеся в настоящее время сведения о прижизненном гемолизе весьма недостаточны, и поэтому приходится пока черпать сведения из опытов гемо-лиза в пробирке, учитывая, конечно, при этом всю относительность их значения для установления истинного прижизненного гемолитического процесса.
Исследования Баркана (Barkan) показали, что в циркулирующей крови внутри эритроцитов происходит расщепление гемоглобина на билирубин, железо и глобин. Билирубин и отщепленное железо проникают через оболочку эритроцита в плазму крови. Здесь образуется комплекс билирубин-альбумин и комплекс железо-глобулин. Баркан считает, что в эритроците наряду с гемоглобином образуется псевдогемоглобин, который характеризуется тем, что его простетическая группа имеет открытое кольцо, согласно формуле:
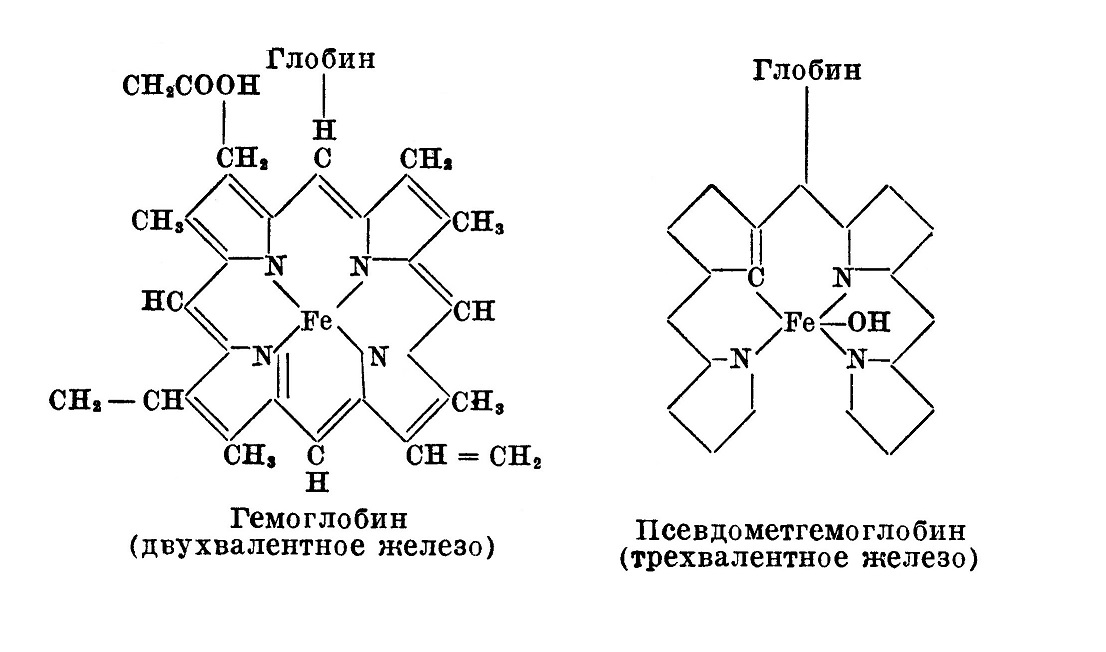
Отщепляя железо, молекула псевдометгемоглобина распадается на глобин + железо + билирубин, согласно формуле:
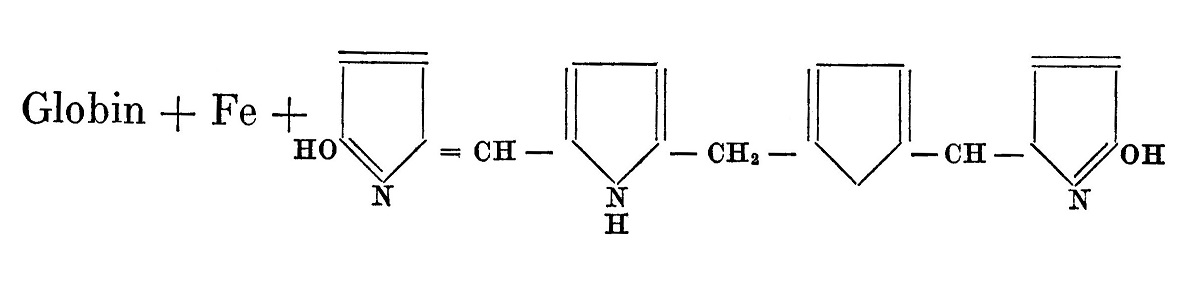
Подсчитывая по количеству отщепленного железа в плазме крови количество распадающегося ежедневно гемоглобина, получаем, что эритроцит в течение своего трехнедельного жизненного существования может потерять до 1/10 гемоглобина. Пересчитывая количество отщепляемого в циркулирующей крови билирубина на образующиеся ежедневно желчные пигменты, также получаем, что оно покрывает самое большее 10% общего количества желчных пигментов. Остаток должен покрываться путем гибели эритроцитов.
Порфирины образуются из простетической группы гемоглобина также путем отщепления железа, но в них кольцо остается замкнутым, что является необходимым условием для получения характерного спектра порфирины.
Тогда как при нормальных условиях простетическая группа гемоглобина почти полностью переходит в билирубин (по номенклатуре Фишера — развернутый гемин III) и только в виде следов выделяется как порфирин, при патологических условиях иногда выделяются большие количества порфирина.
В настоящее время известен целый ряд порфиринов, все они в основном сводятся к порфину:
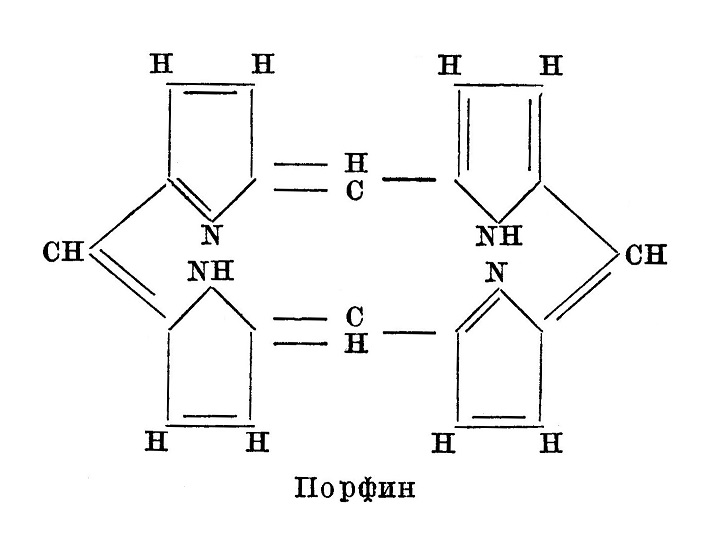
Порфин химически близок гемину, так как производится из гемина путем удаления из последнего железа. Поэтому было бы заманчиво предположить, что порфирины являются промежуточной ступенью при образовании билирубина из гемина.
Фишер считает однако, что образование порфиринов является своего рода браком при пигментном обмене. Выделяемое ежедневно мочой некоторое количество порфиринов состоит главным образом из всосавшихся из кишечника порфиринов, т. е. пищевых порфиринов, образовавшихся большей частью в результате разложения мышечного гемоглобина бактериями — из так называемых экзогенных порфиринов. Эндогенные порфирины образуются и выделяются в небольшом количестве, они представляют собой определенную константную величину, в противоположность экзогенным порфиринам, количество которых зависит от качества вводимой пищи. Оба вида порфиринов могут быть разделены путем установления по возможности бедной порфиринами диеты. Таким образом, можно определить величину эндогенных порфиринов. Возможно, что эндогенные порфирины появляются также в результате неправильного синтеза .гемоглобиновой молекулы,, если синтез прекращается на стадии образования порфирина. В норме эритроциты дают слабый спектр порфиринов (ван ден Берг), а при микроскопическом исследовании при освещении ультрафиолетовыми лучами отдельные эритроциты и нормобласты дают явления характерной флюоресценции — это заставляет предположить, что и при нормальном образовании гемоглобина часть порфиринов остается неиспользованной и находится в молодых эритроцитах. При увеличении эндогенных порфиринов Эппингер говорит о порфиринурии как о болезни и противопоставляет ей экзогенную порфиринурию как симптом, свидетельствующий о вторичном нарушении порфиринового обмена веществ, имеющем место при различных заболеваниях, например, при заболеваниях желудочно-кишечного тракта, при расстройствах кровообращения, при болезнях крови и особенно при заболеваниях печени. Определение количества порфиринов в моче у больного при бедной порфиринами растительной диете до и после нагрузки 20 г кристаллического гемоглобина считается функциональной пробой печени: при нормальной функции печени всосавшиеся из кишечника порфирины должны быть переведены в уробилин и увеличения порфиринов в моче не должно наступить.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 