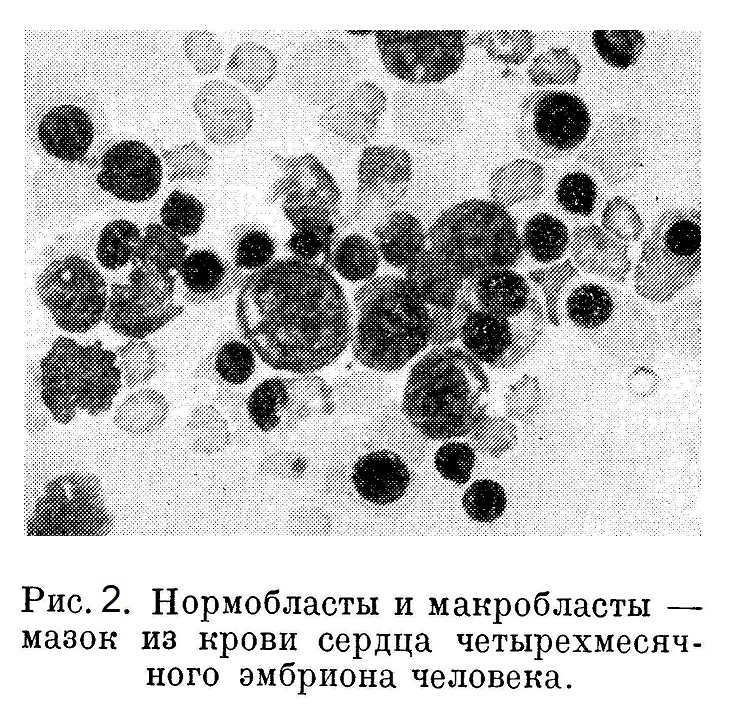
У эмбриона длиной в 2,5 мм закладывается печень. Это происходит путем врастания железистого эпителия duodeni в мезенхимальную ткань. По мере формирования из тяжей эпителиальных клеток и лежащих между ними в мезенхимальной ткани капилляров образуются печеночные дольки. Из мезенхимальных клеток, а также путем деления эндотелия капилляров нарождаются новые мегалобласты. По мере увеличения печени все более и более ослабевает кроветворение во всей мезенхиме зародыша, так что вне печени только кое-где удается обнаружить незначительные кроветворные очажки. Среди эритробластов характера мегалобластов начинают появляться эритробластического типа клетки в 11/2—21/2 раза меньше по объему — нормо- и макробласты. Ядро макробластов больше, чем у мегалобластов второй генерации, богато базихроматином с характерным расположением оксихроматина. Сгущения базихроматина у нормобластов часто дают фигуру колеса.
Нормобластические ядра очень быстро сморщиваются, делаются пикнотичными с густым базихроматином и грубой структурой оксихроматина. Между прочим, по выраженной структуре оксихроматина при альтмановской окраске в моей модификации можно с легкостью отличить эритробласт от какой-либо другой молодой клетки, так как в последней рисунок оксихроматина менее резко выражен. В отношении протоплазмы нормо- и макробласты по своей способности окрашиваться не отличаются от мегалобластов: одни из них полихроматичны, другие ортохроматичны; гемоглобина протоплазма содержит меньше, соответственно меньшему объему клетки (рис. 1, 2 и 3).


Наряду с ядерными появляются и безъядерные эритроциты. Процесс, ведущий к исчезновению ядра, идет через пикноз, сморщивание и распад ядра на отдельные глыбки хроматина. Некоторые ядра рассасываются, не пройдя стадий пикноза, — можно наблюдать, как они как бы тают, постепенно растворяясь в протоплазме. Каким образом исчезают последние остатки ядра, морфологически неясно; возможно, что когда остатки ядра из некробиотического состояния переходят в некротическое, т. е. превращаются в мертвую частицу, они перестают воспринимать краску (как это обычно и бывает при некрозе клеток) и растворяются в протоплазме. Некоторые авторы считают, что ядро выталкивается из клетки. Часто около еще хорошо сохранившегося ядра имеется отшнуровавшееся от него круглое тельце из хроматина, при окраске азур-эозином ярко вишневого цвета. Изредка этих телец бывает несколько. Они часто сохраняются в эритроцитах, лишенных ядра; это тельца Жолли-Хоуэлла (Jolly-Howell).
Тельца Жолли являются, таким образом, отшнуровавшимися частицами, которые труднее рассасываются, чем оставшееся ядро; их поэтому нужно отличать от сегментов распадающегося ядра. После окончательного рассасывания ядра в эритроцитах остаются небольшие остатки его в виде так называемых краевых телец — Randkorperchen. При усиленной регенерации, особенно после удаления селезенки, остатки ядра бывают видны в интенсивно окрашенном азур-эозином препарате в виде мелкой хроматиновой пыльцы. Краевые тельца хроматина не имеют и поэтому не окрашиваются.
В неокрашенном препарате краевые тельца — по одному, реже по 2—3 в эритроците — можно видеть в темном поле зрения как блестящие точки (Шиллинг). При окраске нильблаусульфатом (суправитально) их можно обнаружить в норме от 0 до 3%.
В мегалобластической системе аналогичными нуклеиновым тельцам Жолли являются тельца, также отшнуровавшиеся от ядра, но при окраске азурными смесями окрашивающиеся в голубой цвет (тельца Жолли — вишневокрасные, азурофильные). Мне кажется правильным назвать эти тельца парануклеиновыми — от парануклеина, ядра, также окрашивающегося в голубой цвет. Парануклеиновые тельца в мегалобластах имеются в большом количестве в крови эмбрионов человека от 1,5 до 2 см длины, также местами и в мегалоцитах. При витальной окраске они резко окрашиваются в темно-синий цвет, так что очень легко проследить весь процесс отшнуровывания их от поверхности ядра.
Эритроциты, получившиеся из мегалобластов, такой же величины, как и последние, — мегалоциты; из нормобластов получаются нормоциты; переход мегалобласта путем деления в нормобласт не установлен.
В мазках из печени и в крови из сердца человеческого эмбриона длиной в 2 см встречаются эритроциты с фуксинофильной зернистостью (по альтмановскому способу); несомненно это есть сохранившаяся зернистость мегалобластов, resp. нормобластов.
В большом количестве мне пришлось прежде, когда я впервые описала эту зернистость, видеть ее в эритроцитах очень молодых эмбрионов мыши.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
| 