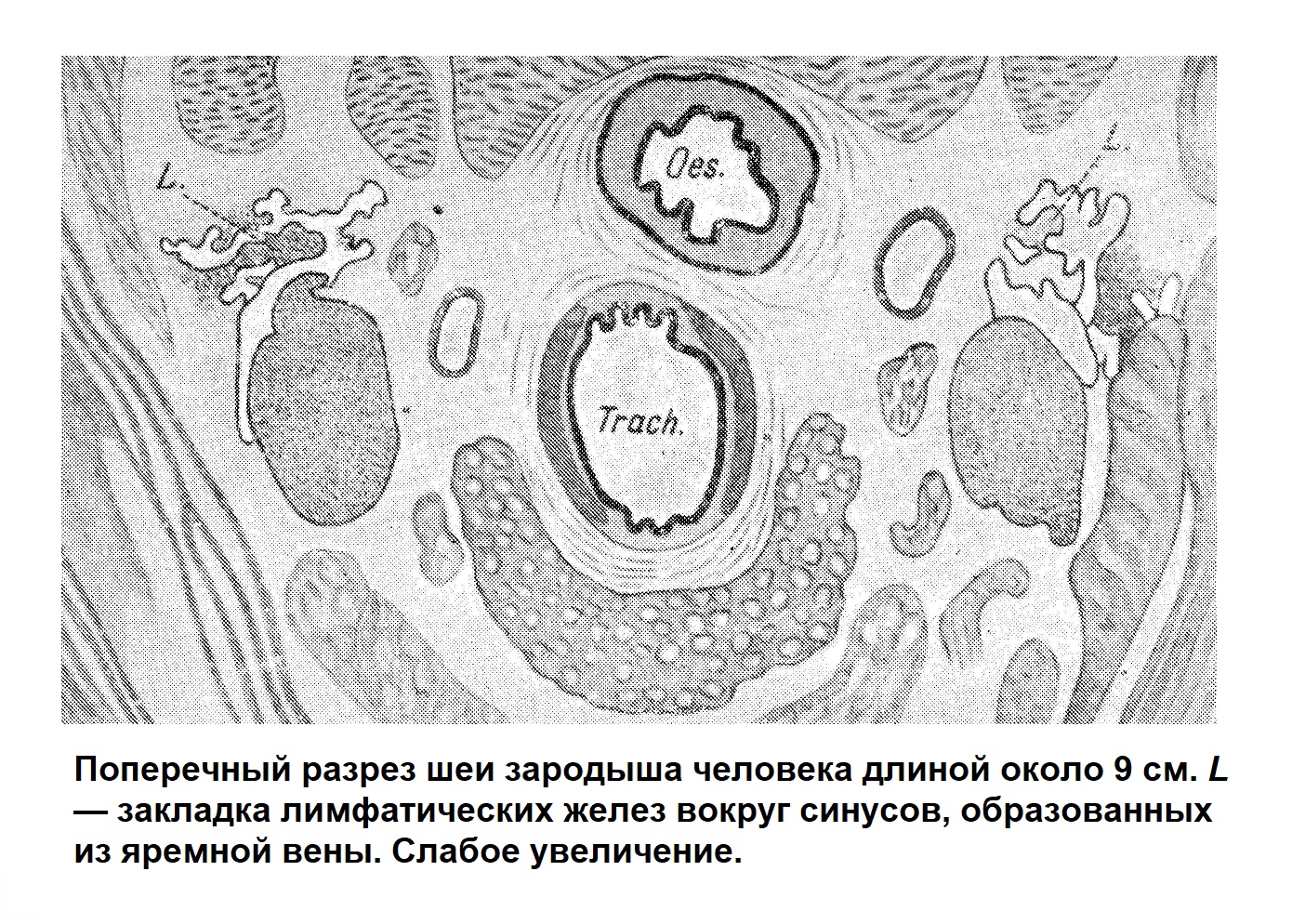
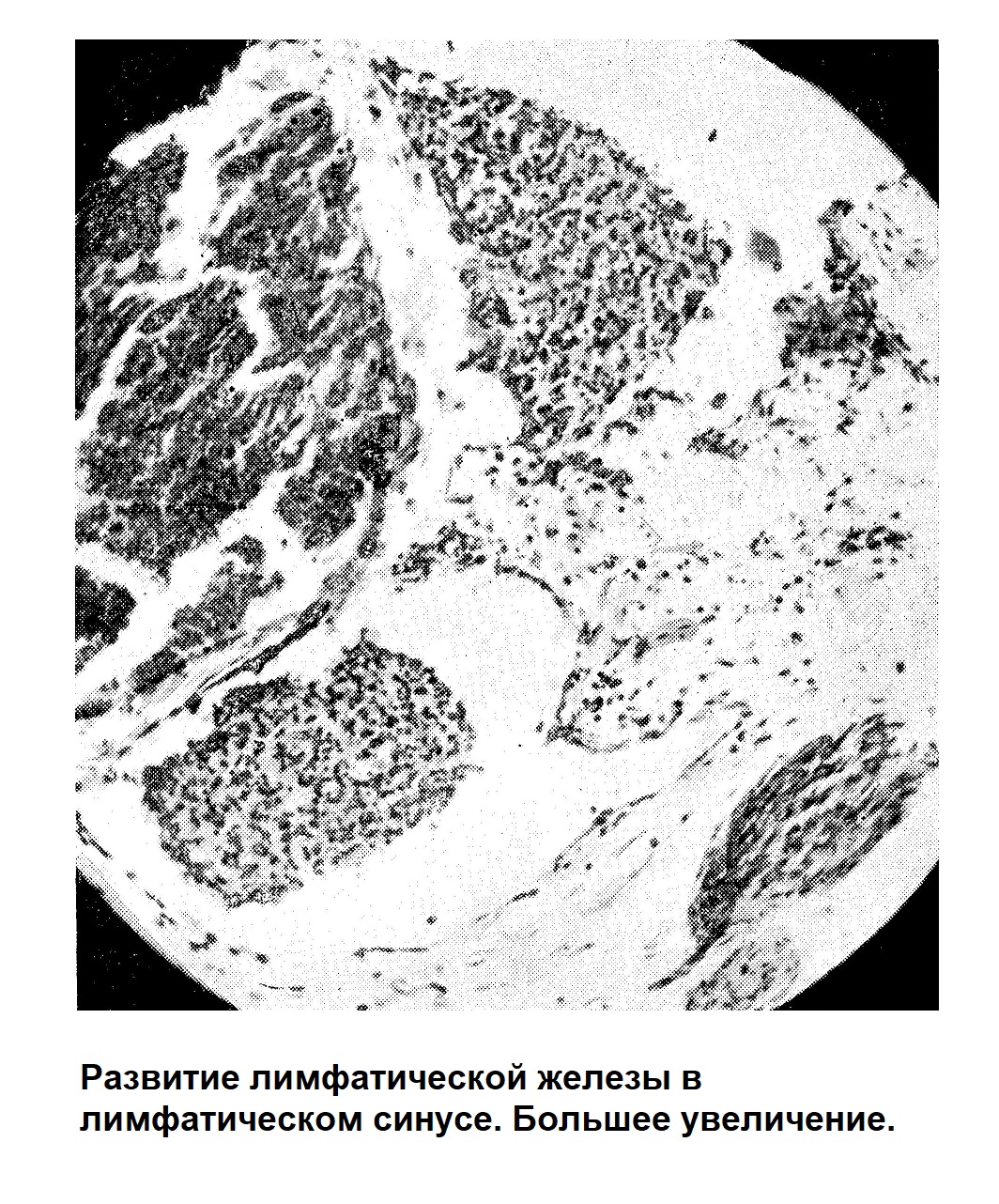
Закладкой лимфатических желез является [Кларк (Clark), Сабин] образование лимфатического синуса около больших вен (яремная, полая вена и др.). Формирование его очень легко проследить на шее, на уровне щитовидной железы. У эмбриона человека длиной в 4,5—5 см получается сначала небольшое, а затем все более увеличивающееся дивертикулообразное выпячивание эндотелия яремной вены. Таким образом, получается сосуд (лимфатический синус), в который сначала попадает кровь из вены, а потом он совершенно отшнуровывается и содержит только лимфу. По периферии этого лимфатического синуса (рис) появляются скопления клеточных элементов вокруг мелких кровеносных сосудов и железе у человеческого эмбриона длиной в 13 см еще преобладают, миэлогенные клетки; несколько позже большинство клеток формирующейся лимфатической железы лимфоидного характера.
Итак, у двухмесячного эмбриона человека, когда уже все виды лейкоцитов циркулируют в крови, лимфоциты еще отсутствуют: мы не имеем ни закладки лимфатических органов, ни отдельных лимфобластических очагов в мезенхиме; даже в местах закладки лимфатических органов на ранних стадиях развития обнаруживаются только миэлогенные элементы. Из этого мы должны сделать заключение, что, во-первых, миэлогенные элементы являются онтогенетически более ранними, развиваются непосредственно из мезенхимальных клеток, во-вторых, что в тех же самых местах, при изменившемся только раздражителе, мезенхимальная ткань продуцирует другие клетки, онтогенетически более поздние — лимфоциты.
Лимфоцит развивается или непосредственно из мезенхимальной или ретикуло-эндотелиалъной клетки, ил снчала появляется большая клетка с базофильной протоплазмой и большим, бедным базихроматином ядром с ядрышками — лимфобласт. Базихроматиновая сетка его ядра несколько отличается от сетки ядра миэлобласта, однако в некоторых случаях диференцировать чрезвычайно трудно, даже невозможно. Фуксинофильная зернистость протоплазмы при альтмановской окраске, расположенная вокруг ядра, хорошо выражена; такого же вида зернистость имеется и у зрелых лимфоцитов — шридовская зернистость. Но, как мы уже видели, и у миэлобластов имеются такие же зерна, так что диференцировать эти клетки друг от друга по фуксинофильной зернистости невозможно.
Негели считает, что фуксинофилъная зернистость лимфоцитов и миэлобластов по существу совершенно различные образования; это, конечно, вполне допустимо; однако и те, и другие дают характерную митохондральную окраску янусгрюном.
Неоспоримой остается стабильная оксидазная реакция: у лимфобластов она всегда отрицательная, у некоторых миэлобластов она положительная, и в этих случаях различие между ними очень резкое. Лабильная оксидазная реакция у некоторых лимфоцитов изредка может быть также положительной. Ядро молодого лимфоцита при созревании быстро темнеет, клетка переходит в обыкновенный малый лимфоцит; этот последний имеет характерное темное ядро с густым базихроматином, в котором совершенно не видна при обычных окрасках нежная структура оксихроматина. Вокруг ядра у одних лимфоцитов — узкая кайма голубоватой протоплазмы, у других она значительно больше, и ядро тогда расположено несколько эксцентрично, так что величина клетки вариирует. Около ядра в протоплазме имеется оксифильная зона с центросомой в середине. При окраске азурсодержащими смесями в протоплазме обнаруживаются (однако не всегда) азурофильные зерна различной величины, кругловатой формы; нужно думать, что это продукт межуточного липоидного обмена веществ лимфоцита. В то время как Тюрк, Иокомори (Turk, Jokomori) и др. устанавливают участие лимфоцитов в липоидном обмене, Ашоф и Рент (Resch) его совершенно отрицают.
В циркулирующей эмбриональной крови незрелые лимфоциты встречаются редко; некоторые авторы подразделяют зрелые лимфоциты по величине их протоплазмы на малые, средние и большие, но это вряд ли имеет какое-нибудь практическое значение.
По мере развития закладки лимфатической железы изменяется не только состав окружающих лимфатический синус клеток — миэлогенные элементы все более замещаются лимфоидными, но и самая основа, в которой развивается кроветворная ткань, начинает терять свой характер недиференцированной мезенхимальной ткани, становится петлистой, ретикулярной. Стенка петель состоит из узких протоплазматических прослоек с утолщением в тех местах, где находится ядро. По установившемуся взгляду эта ретикулярная ткань состоит не из отдельных, соединенных между собой клеток, а представляет собой синцитий, т. е. ткань, в которой ядра делятся без последующего деления протоплазмы. Таким образом, получается сплошная протоплазматическая сеть с заложенными в ней ядрами. Как эндотелиальные клетки, происшедшие из мезенхимальных, частично сохраняют способность развиваться в кровяные клетки, точно так же и ретикулярная ткань сохраняет способность под влиянием различных раздражителей давать различные кровяные клетки.
Таким образом, мы видим, что уже у очень малых эмбрионов заложена ретикуло-эндотелиальная система, которая также рано начинает выявлять крайне разнообразную функцию, свойственную ей и во внеутробной жизни. Уже на втором месяце эмбриональной жизни погибают в большом количестве красные кровяные тельца; в мазках из печени мы находим в изобилии эндотелиальные клетки с ядерными и безъядерными эритроцитами в протоплазме и отложением кровяного пигмента, т. е. явление фагоцитоза. Эндотелий пожирает отмирающие форменные элементы (красные и белые кровяные тельца), — таким образом образуются тканевые фагоциты, или гистиофагоциты. Фагоцитарная функция свойственна всей ретикуло-эндотелиальной системе, однако в различной степени. В этом направлении происходит также дальнейшая диференциация в том смысле, что по всей системе выделяются отдельные клетки, собственно фагоциты, которые в первую очередь пожирают все шлаки, циркулирующие в крови. В печени такими клетками являются так называемые купферовские клетки, происшедшие из эндотелия капилляров печени; они имеют звездчатую форму, вдаются в просвет сосуда, жадно поглощают форменные элементы и перерабатывают гемоглобин с образованием железа и билирубина.
В мазках крови, взятых из сердца, обычно встречаются пласты слушанного эпителия (артефакт); в них нередко можно также наблюдать фагоцитированные эритроциты, как это видно из схемы. Из этого можно заключить, что по мере появления в крови большого количества шлаков сенсибилизируется в смысле фагоцитоза и эндотелий больших сосудов (то же явление мы встречаем при патологии у взрослого организма).
Несомненно, что ретикуло-эндотелиалъные клетки участвуют в гемоглобиновом обмене не только тем, что поглощают эритроциты и кровяной пигмент, но и тем, что скопляющееся в них железо постоянно отдается обратно в кровь и идет вновь на выработку гемоглобина; возможно, что и пиролловые основания, являющиеся конечным продуктом распада гемоглобина, вновь поступают в кровообращение и путем синтеза дают гемоглобин. Где происходит образование гемоглобина — в самой ли эритробластической клетке или же имеются еще межуточное синтетические станции, нам еще неизвестно.
Перед нами встает также вопрос: когда начинают вырабатываться желчь, билирубин, которые являются продуктом гемоглобинового распада? Возможно, что на ранней стадии развития зародыша расщепление гемоглобина происходит вплоть до билирубина в ретикуло-эндотелии, откуда билирубин поступает в кровь. По мнению Ашофа, билирубин и во взрослом организме образуется в ретикуло-эндотелиальной системе, а печень является только выводящим органом.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
|
