Повсеместно ли в мезенхиме эмбриона самостоятельно образуются кровяные островки и из них уже кровеносные сосуды с кровяными клетками или же васкуляризация идет исключительно из area vasculosa, — этот вопрос не получил еще окончательного решения. Во всяком случае на этой стадии развития кроветворная функция всецело выполняется сосудистыми образованиями. Мы уже указывали, что клетки, выстилающие просвет сосудов, еще мало дифференцированы, — они по структуре своей больше напоминают недифференцированную мезенхимальную клетку, чем эндотелий. В дальнейшем эти клетки, размножаясь, дают небольшие утолщения стенки сосуда, затем отделяются и наполняют кровяное русло новыми первичными кровяными элементами. Последние очень скоро начинают видоизменяться, в их протоплазме начинает появляться гемоглобин; при окраске протоплазма из голубой становится лиловатой, полихроматофильной, т. е. окрашивается и кислой, и щелочной краской. По величине клетки продолжают оставаться очень большими, ядро также большое, бедное базихроматином, структура оксихроматина плохо выражена. Таким образом, первичные кровяные клетки превратились в большие ядросодержащие красные кровяные тельца — мегалобласты. Мегалобласты некоторое время являются единственными клетками в циркулирующей крови. Эндотелий сосудов, размножаясь, продолжает продуцировать мегалобласты. Наряду с мегалобластами, происходящими эндоваскулярно из недиференцированного эндотелия, можно всюду обнаружить также мегалобласты, расположенные в виде муфты вокруг кровеносных сосудов; здесь они образуются из прилегающих снаружи к сосудам мезенхимальных клеток, которые всюду сопровождают сосуд, составляя как бы часть его стенки.
Мезенхимальные клетки и у взрослого организма остаются постоянными спутниками кровеносных сосудов. Это дало повод отдельным авторам называть эти клетки перителием. Перителиальные клетки имеют амебоидную форму, овальное ядро; однотипные клетки находятся повсюду в соединительной ткани — так называемые клазматоциты, полибласты. Все эти клетки легко выявляются при введении животному различных красок, например, пирроловой синьки (поэтому их также называют пирроловыми клетками), трипановой синьки и др.
Таким образом, являясь дериватами мезенхимальных элементов, эти клетки представляют собой строго диференццрованные клетки, входящие в состав обширной ретикулоэндотелиалъной системы. Они проявляют, однако, в норме свою кроветворную способность только в кроветворных органах, так как в остальных тканях организма для них нет соответствующего функционального раздражителя. При различных же воспалительных, опухолевых и других процессах периваскулярная мезенхима выявляет свою потенциальную способность образовывать кровяные элементы.
Наряду с мегалобластами, имеющими большое светлое сетчатое ядро и полихроматофильную протоплазму (мегалобласты первой генерации), вскоре появляются клетки такой же величины, даже местами несколько большие, с богатой гемоглобином резко оксифильной протоплазмой, однако с совершенно иным ядром — темным (богатым базихроматином), небольших размеров, местами с выраженным расположением базихроматина в виде колесика — мегалобласты второй генерации. Возможно, что часть этих мегалобластов произошла, как указывает Негели (Naegeli), из мегалобластов с большим светлым ядром путем сморщивания; однако через очень короткий промежуток времени мегалобласты с большим светлым ядром, с характерной структурой совершенно исчезают не только из циркулирующей крови, но они не обнаруживаются и в местах их образования (рис. 1 и 2).
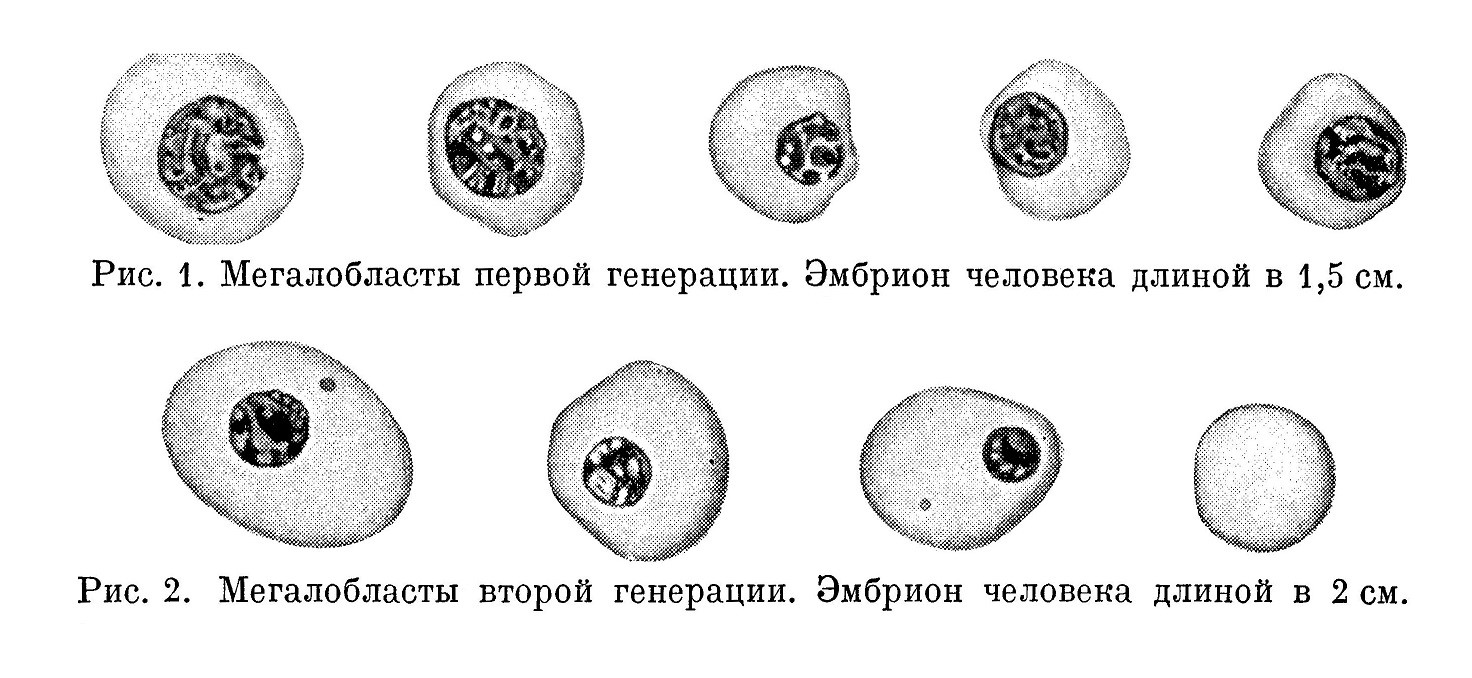
Таким образом, мы видим, что клетки кровяных островков постепенно развиваются в ядросодержащие красные кровяные тельца — мегалобласты; иными словами, клетки кровяных островков являются первичными эритробластами.
Мезенхимальные клетки, лежащие в центре кровяных островков, дифференцируются в эритрогонии или в проэритробласты, дальнейшее развитие которых зависит от нейрогуморального фактора. Эритрогонии или проэритробласты первого и последующего типа кроветворения отличаются друг от друга только величиной. Ни в протоплазме, ни в ядре никаких отличительных признаков не имеется. Различие появляется только в момент диференциации проэритробласта в мегалобласт. Мегалобласт по структуре ядра и по форме протоплазмы (более или менее овальной) представляет собой характерную клетку, отличную от других эритробластических элементов. Поэтому большой величины проэритробласт (или эритрогония) не определяет собой мегалобластического типа кроветворения.
Некоторые авторы все же признают особую форму — промегалобласт; однако по этому поводу мы читаем в монографии Генинга и Кайльгаки (Henning и Keilhacki): «Сходство между проэритробластом и промегалобластом несомненно значительно большее, чем между проэритробластом и миэлоб ластом».
Уже на первых стадиях развития кровяных клеток мы видим, что они не происходят непосредственно из недиференцированнцх клеток мезенхимы, но что последние сначала переходят в эндотелиального и перителиального характера клеточные элементы. На всем протяжении зародышевого периода развития, а также и в постэмбриональном периоде дериваты мезенхимальной ткани сначала в виде эндотелия, а затем, как мы увидим ниже, и ретикулярного синцития в большей или меньшей степени участвуют в кроветворении. В понимании механизма развития кроветворного процесса до сих пор нет единого мнения. Основное противоречие различных школ сводится главным образом к следующему: идентично ли эмбриональное и поетэмбрионалъное кроветворение? Негели говорит — нет, не идентично: у эмбриона кровяные клетки нарождаются из мезенхимальных клеток, а в постэмбриональном периоде происходит только созревание уже имеющихся клеток с определившейся в своем потенциальном развитии направленностью. Это имеет место при нормальном, здоровом состоянии организма. При патологии оживают дремлющие мезенхимальные клетки и снова начинают кроветворить по типу различных стадий эмбрионального кроветворения — это кроветворение характера метаплазии. Согласно этому пониманию, если в постэмбриональной жизни появляются мегалобласты, родоначальников которых в данном периоде не существует в кроветворной ткани, следовательно, здесь должно иметь место появление реактивированной мезенхимальной ткани с потенциями эмбрионального периода. Имеем ли мы морфологические доказательства этому положению ? Нет, никакими морфологическими методами не удается доказать наличие резервной, недиференцированной вездесущей мезенхимальной ткани. Да и само понятие эмбриогенизации в смысле возврата к эмбриональному состоянию требует доказательства своего права на существование. Мы знаем, что зародышевая клетка на ранних ступенях развития обладает большой потенцией к развитию; по мере диференциации она теряет способность многостороннего развития и этой способности никогда больше приобрести не может. Если у диференцированной клетки упрощаются структуры, клетка обезличивается; это идет не за счет накопления потенций, а за счет их снижения. Поэтому наша точка зрения такова: мезенхимальная ткань диференцируетея в эндотелиальную, resp. перителиальную, ткань, затем в ретикуло-эндотелиальную, которая сохраняет свои потенциальные возможности в течение всей жизни организма. Дальнейшая ее диференциация в смысле образования тех или иных кровяных элементов происходит в зависимости от функциональных раздражений, которые она получает. Раздражения идут главным образом через посредство нервных связей, которые осуществляются богатой сетью нервных волокон, пронизывающих эндотелий (а позднее и ретикулярный синцитий). Первичная, непосредственная раздражимость клеточной протоплазмы является ведущим фактором главным образом в отношении фагоцитарной функции клеток. Таким образом, мы считаем, что, например, мегалобластический характер эритропоэза как в эмбриональном, так и в постэмбрионалъном периоде жизни организма обусловливается не особенностями кроветворной тйани, а изменением характера раздражителя. Так, мы знаем, что мегалобласты при пернициозной анемии появляются в зависимости от отсутствия нормальной секреции слизистой оболочки желудка, так называемого внутреннего фактора Кэстла (Castle), фактора, по-видимому, природы фермента. Возможно, что на ранних стадиях у зародыша также отсутствует секреция слизистой оболочки желудка и соответственно этому образование эритробластов идет по мегалобластическому типу.
В периоде эмбрионального развития, а постнатально при различных патологических процессах приходится наблюдать значительные отклонения от средней величины клеток. Это особенно хорошо выступает на кривых Прайс-Джонса при мегалобластическом кроветворении. При развитии мегалобластов из эритрогоний особенно резко выступает укрупнение клетки при диференциации. «Рост клетки сопровождает специализацию при остановке деления, — говорит Вермель, — кроме того, надо принимать во внимание, что изменчивость размеров клеток состоит в прямой связи с их специфической функцией». Этим объясняется, например, что нейробласты мелки, а развивающиеся из них двигательные ганглиозные клетки чрезвычайно крупны, что кроветворная клетка может быть мелкой, а развивающаяся из нее клетка крови может стать очень крупной. Согласно этой теории, организм как целое оказывает на клетки ограничивающее рост влияние. Это координирующее влияние организма ослабевает в отношении клеток злокачественных новообразований; ослабления его не удается, однако, выявить, исходя из данных, полученных методом вариационной статистики при лейкемии.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
|
